
Autor/es: Aníbal J. Pordomingo EEA “Guillermo Covas” INTA Anguil Facultad de Ciencias Veterinarias UNLPam 2013
La composición del alimento a utilizar es el componente central de la definición del costo. Las dietas pueden variar en su grado de complejidad yendo de las más simples que sólo son ingredientes utilizados como ingresan al campo y solamente mezclados con un mixer, hasta aquellas en las que el productor procesa los ingredientes (comúnmente los granos) y compone su propio núcleo vitamínico y mineral.
La mayoría de los planteos actuales en la región prefieren las mezclas simples donde en el campo se mezcla el grano (molido, aplastado o entero) con un concentrado proteico (concentrados comerciales, o subproductos de la industria aceitera como la harina de girasol, de soja o de semilla de algodón), (que a su vez aportan fibra), con un núcleo vitamínico y mineral que provee los macro y micro minerales, a los que se les suma una fuente de fibra larga en forma de rollo, heno (ofrecido entero, desmenuzado o molido) o en forma de ensilajes. Frecuentemente se agre- ga urea para aportar nitrógeno, promover la producción de proteina ruminal y reducir la necesi- dad de harinas proteicas.
En el Cuadro 1 pueden observarse algunas dietas comunes en planteos de engorde de terneros (T1 a T5) y de novillos en terminación (N1 a N3) en la región central de Argentina. Estas dietas son simples, propias de una región con escasa diversidad de sub-productos de natu- raleza proteica. Puede observarse que, aunque variables en sus componentes, son muy similares en composición nutritiva dentro de cada categoría.
En el Cuadro 2 se resume a manera de referencia la composición proximal y de oferta energética de los principales alimentos utilizados en la conformación de dietas de corral. Los valores consignados son provienen de promedios de determinaciones realizadas sobre muestas recibidas en los laboratorios de INTA y referencias bibliográficas. La utilización de los datos presentados en el cuadro deben ser tomados como orientativos pero para la conformación y ajuste de dietas se deberá proceder al análisis de laboratorio de los insumos debido a la gran variabi- lidad de la composición de los alimentos. En particular deben analizarse los subproductos agroindustriales húmedos como las burlandas, el gluten feed, las harinas proteicas y los silajes.
El grano
El grano es el componente mayoritario en las dietas de feedlot clásico, comúnmente ex- cede el 65% del total del alimento y define la oferta de energía metabolizable y las características físicas del alimento. El tipo de grano (McCollough et al., 1972; Perry, 1976) y el procesado o la presentación (Rooney y Pflugfelder, 1986; Stock et al., 1987 ab) definen el grado de aprovecha- miento. El rumen es, aún en el feedlot con dietas de alto energía (ricas en almidón), el sitio prin- cipal de utilización del grano. La fermentación ruminal es el proceso fundamental para generar la energía necesaria para el crecimiento y engorde (Owens et al., 1997). En ese sitio se fermenta la mayor parte de la fracción digestible del grano (60 al 85%) y la mayor proporción del almidón (más del 90%) si ha sido expuesto al licor ruminal (Owens et al., 1997, Huck et al., 1998; Philip- peau et al, 1999 a,b; Cooper et al., 2002b).
El grano es fermentado por la flora ruminal, la que obtiene del mismo la energía y nu- trientes para su proliferación. La masa microbiana fluye permanentemente desde el rumen hacia el estómago verdadero donde es digerida (digestión ácida). Por ser rica en proteína microbiana su mayor aporte a la nutrición del animal es en la forma de aminoácidos y péptidos, los que son absorbidos en el intestino. La proteína que aporta esa masa microbiana es generalmente de ma- yor valor biológico (más balanceada y completa para los requerimientos del animal) que la pro- teína ingerida (Huntington, 1997).
En las dietas de engorde a corral comunes en Argentina la calidad de esa proteína que al- canza el intestino delgado, si se han formulado para cubrir los requerimientos de proteína meta- bolizable, es suficiente para utilizar la energía metabolizable que tales dietas ofrecen. La mejora de eficiencias por incorporar proteínas de baja degradabilidad ruminal es inexistente o escasa (Pordomingo et al., 2003).
La síntesis de proteína microbiana a partir de fuentes de nitrógeno no proteico es uno de los elementos centrales de la alimentación a corral de bajo costo, donde la proteína bruta de la dieta es de calidad nutritiva muy inferior a la que llega al intestino del bovino. La masa bacteria- na es también proveedora de lípidos esenciales -que el animal no puede sintetizar en su metabo- lismo- y minerales, tomados del alimento pero asociados a proteína bacteriana.
La fermentación ruminal normal ocurre en ausencia de oxígeno. En un medio carente de oxígeno, el proceso degradativo libera energía pero no termina en la formación de agua como producto final sino en la generación de ácidos grasos volátiles. Estos metabolitos son considera- dos residuales de la fermentación pero son todavía energéticamente ricos, siendo los más impor- tantes el acético, el propiónico y el butírico. También se generan otros ácidos de metabolismo intermedio entre los que debe citarse el ácido láctico (Church, 1988).
De los ácidos grasos principales citados, el acético es el principal precursor de la síntesis hepática de lípidos, componentes de los triglicéridos, y el propiónico es el principal precursor para la generación de glucosa (gluconeogénesis) (Smith y Crouse, 1984). El ácido láctico es más fuerte que los otros y de remoción lenta del medio ruminal. Su acumulación incrementa rápida- mente la acidez del rumen –acidifica el medio, torna la absorción de ácidos más lenta y puede generar acidosis, parálisis o coma muscular y el síndrome del empacho.
En las dietas de alto grano (fermemtación preferentemetne amilolítica), el pH ruminal oscila entre 5.0 y 6.0. Luego 2 horas del suministro de alimento o entre 1 y 2 horas de ocurrida la ingesta, se han descripto depresiones del l pH ruminal a valores de 4.5, pero 1 hora más tarde debería ubicarse en valoresde 5 o superiores (Pordomingo et al., 1999b) Esta dinámica de la acides se encuentra regulada con la capacidad buffer de la saliva y el proceso de remosión de ácidos desde el rumen. Si el pH se mantiene por debajo de 5 durante períodos prolongados, mas de 3 horas, la erosión de epitelios, laminitis y degeneración de papilas comienza, aumentan los riesgos de inflamación, abración y lastimado de la dermis con ulceraciones consecuentes e infec- ción.
El intestino delgado es otro sitio de utilización del almidón (Owens et al., 1986). Esa di- gestión complementaria que puede ocurrir a nivel de tracto digestivo inferior (intestino delgado) hace que la digestibilidad del almidón sea casi completa (92 al 99%) (Waldo, 1973; Philippeau et al., 1999a; Pordomingo at al., 2002). Pero, el tipo de almidón y el procesado del grano tienen una influencia directa sobre la tasa y sitio de digestión (Galyean et al., 1976; Nocek y Tamminga, 1991). Los granos de trigo y cebada poseen un almidón de mayor degradabilidad ruminal que el sorgo y el maíz (Boeto y Melo, 1990; Herrera-Saldana et al., 1990; Waldo, 1973). Estos últimos tienen una estructura proteica entre gránulos que impide la rápida exposición al licor ruminal y retarda el ataque microbiano (Rooney y Pflugfelder, 1986). Entre los maíces podrían citarse diferencias similares al comparar dentados, semi-dentados y duros. Philippeau et al. (1999a) reportó rangos 40.6 a 77.6% de fermentabilidad ruminal del almidón, con los maíces vítreos tipo flint en el extremo inferior y los más harinosos tipo dent en el superior. Similares resultados fue- ron obtenidos por Maresca et al. (2002).
Existe siempre, en mayor o menor medida, una fracción de almidón que escapa a la fer- mentación ruminal y que alcanza el intestino delgado donde puede ser digerida enzimáticamente y absorbida directamente como monómeros o dímeros de glucosa. El escape de almidón hacia el intestino delgado puede incluso mejorar la eficiencia de utilización de la dieta por su digestión directa (Waldo, 1973; Philippeau et al., 1999a), afectar el sitio de digestión del nitrógeno, y me- jorar la eficiencia de conversión de la materia seca (Russell et al., 1981; Streeter et al., 1989). Este mecanismo, sin embargo, es limitado porque la capacidad intestinal para remover almidón de la ingesta es inferior a la de los monogástricos (Huck et al., 1998; Owens et al., 1986; Russell et al., 1981; Swanson et al., 2002).
La capacidad del intestino delgado de digerir almidón no debe ser sobre-estimada, el es- cape de almidones hacia el intestino grueso promueve la fermentación amilolítica en ese sitio, pero sin posibilidades de remoción de los ácidos grasos volátiles (productos de fermentación) via absorción por la mucosa intestinal. En otras especies esta capacidad está desarrollada (caballos, conejos) pero no así en rumiantes. Solo se absorven en intestino delgado el agua y algunos elec- trolitos pero no moléculas orgánicas. Esa fermentación genera además importantes cantidades de metano e inchado por gas (distensión) del colon.
La acidificación y distensión del intestino grueso, expuesto a una microflora amilolítica (agresiva) genera imflamación de epitelios, basodilatación, sangrado, erosión y descamado del intestino con disrrupción de la re-absorción de agua (diarrea) y pérdida de electrolitos. En ese escenario, se observan heces líquidas, sanguinolentas (sangre fresca, roja) con epitelio intestinal, frecuentemente claras, amarillas o grices muy claras (si se alimentó con harina de girasol). En heces de algunas horas se puede observar la formación de burbujas. En ese caso es evidente que existe un sustrato de fácil y rápida degradación (almidón) que inicia un proceso de putrefacción con generación de dióxido de carbono, óxidos de azufre y otros gases. Esos gases son atrapados en burbujas que se comienzan a proliferar en la superficie de la deposición.
El procesado de los granos mejora la digestibilidad de la materia seca (del grano) y del almidón (Brenttheurer, 1986), incrementa la tasa de pasaje de la ingesta a lo largo del tracto digestivo (Galyean et al., 1976; McNeill et al., 1976; Theurer, 1986; Huck et al., 1998; Brown et al., 2000b), y mejora el aprovechamiento, en particular de granos pequeños o duros (Perry, 1976; Ewing et al., 1986; Philippeau et al., 1999a). Las formas de procesado son diversas y tienen resultados diferentes según se trate de granos secos o húmedos.
Granos secos
La molienda, el micronizado y el aplastado mejoran la exposición ruminal del almidón en cada tipo de grano (Cooper et al., 2002a,b; Brenttheurer, 1986). El cambio es más significati- vo en los de endosperma córneo (duros), pero en los de tipo dent (harinosos) la exposición alcanza los valores máximos (Chandrashekar y Kirleis, 1988; Philippeau et al., 1999a,b).
La forma de molido en polvo es de rápida exposición del almidón y degradación comple- ta. Pero, debido a esa rápida exposición, el molido fino es más susceptible de generar acidez ruminal y acidosis, comparado con otras formas. Además, el polvo de granos es fácilmente inha- lado y puede provocar infecciones respiratorias. En el caso de los granos vestidos (avena y ceba- da) el molido también reduce el tamaño de las fracciones fibrosas externas de protección (glu- mas) a partículas muy finas que por su textura son muy irritantes de epitelios del tracto respirato- rio. Este es un motivo por el cual no se recomienda el molido de granos vestidos. El grano de sorgo y los maíces tipo flint (duros, con endosperma córneo) son los que mejor se adaptan a esta forma de procesado. En el caso del sorgo, genera mejores resultados en conversión que las for- mas partido o entero.
Para reducir los defectos de la forma molida y lograr buena digestibilidad del grano se desarrollaron otros métodos como el micronizado en húmedo, el aplastado (rolado) en seco y el procesado en copos (flakes; grano aplastado, prensado, con humedad y temperatura). Este méto- do transforma al grano en una lámina fina con humedad y produce una gelatinización parcial del almidón. El tamaño de las partículas (láminas, copos o flakes) es grande y las láminas no supe- ran 1 a 1,3 mm de espesor. La digestibilidad es muy alta y no se genera polvo que pueda provo- car complicaciones respiratorias. Por este método se aporta volumen al alimento al expandir el grano en láminas o “flakes”, se logra un consumo homogéneo, alta palatabilidad y baja inciden- cia de trastornos digestivos. El contacto con el licor ruminal y la fermentación es más homogé- neo que en el caso del grano molido (Owens et al., 1997; Swingle et al., 1999). Todos estos atri- butos hacen posible la formulación de dietas de la mayor concentración energética (más de 85% de grano; Owens et al., 1997). Adicionalmente, sobre esos copos (flakes) se absorben o adhieren bien los aditivos líquidos o polvos (sales minerales, vitaminas, ionóforos, etc.). Sin embargo, esta forma de procesado, muy utilizada en EEUU y Canadá, no es común en Argentina por el costo de la inversión en la infraestructura para producir “flakes” y el costo de la energía que in- sume el proceso.
Otros métodos menos eficientes desde el punto de vista físico pero menos costosos en energía son el aplastado en seco o rolado y el aplastado en húmedo (grano conservado en húme- do con 28 a 30% de humedad - o con humedad reconstituida = agregada). Los feedlots que en Argentina que no usan grano de maíz entero han adoptado alguno de estos métodos. El aplastado o quebrado del grano seco es el más común. La exposición del almidón es mayor que la de los granos ofrecidos enteros, especialmente en el caso del grano de sorgo y maíz el tipo duro Philip- peau et al, 1999a). Sin embargo, las diferencias en eficiencia de conversión de estos procesados versus el maíz entero son variables y dependen del tipo de grano de maíz (Philippeau et al., 1999b; Maresca et al., 2002). Si el maíz es de grano grande, dentado o semi-dentado, las dife- rencias entre procesarlo o no son bajas y no se justifican económicamente. Si es del tipo duro y tamaño chico, el procesado sería conveniente.
En el caso del sorgo el procesado es indispensable (McCollough y Brent, 1972; McNeill et al., 1976; Pordomingo y Juan, 2000). En cereales de invierno del tipo vestido (como la avena y la cebada), el aplastado (sin quebrar o moler) mejora la homogeneidad del consumo y de la fermentación, pero la diferencia en conversión con respecto al uso de sus formas enteras es baja o nula (Morgan y Camping, 1978; Owens et al., 1997; Philippeau et al., 1999a; Wang et al., 2003). Elizalde et al. (2003b) encontraron diferencias a favor de la cebada entera versus la aplas- tada en novillitos, alcanzando 1,5 vs 1,3 kg de aumento de peso y conversión similares (6:1) (Cuadro 7).
Beauchemin et al. (2001) reportaron altas digestibilidades del grano de cebada presenta- do en forma molida o aplastada, con un tendencias a mayor grado de digestión ruminal para las formas de procesado intenso (ya sea molido o aplastado), pero las diferencias no resultaron signi- ficativas (Cuadro 8). Por su parte el molido redujo la digestibilidad de la fibra (FDA) aportada por el grano, con respecto a la forma aplastada (muy baja de por sí) (Cuadro 9). La dieta base fue de alto grano, compuesta de 86% de cebada aplastada o molida (Cuadro 10). Estos elementos indican que el procesado puede asegurar la buena utilización de esos granos, pero no necesaria- mente mejorarla.
Los cuadros 3, 4 y 5 resumen resultados de una experiencia reportada por Owens et al. (1997) sobre consumos, aumentos de peso y eficiencias de conversión de engordes de novillos en dietas de alta energía basadas en distintos granos y procesados de los mismos. El procesado en copos y el extrusado favorecieron a algunos granos y no necesariamente a todos en los aumentos de peso o en conversión. Se destaca la performance del maíz entero que se asemejó a las formas extrusada y en copos. Debe mencionarse que los maíces incluidos en estos estudios fueron del tipo dent (dentado grande). De la combinación de efectos, la forma de procesado en copos maximizó la oferta de energía metabolizable de los granos, particularmente del grano de sorgo (Cuadro 7).
Maíz entero
En Argentina se ha difundido el uso del grano entero de maíz por la reducción de costos que implica no procesar el grano. Las eficiencias de conversión logradas en planteos comercia- les con grano de buena calidad (buen tamaño) son similares o escasamente peores (8 a 10%) a las logradas con grano molido o aplastado seco, aún en categorías de buen tamaño corporal (novi- llos). En el mismo ensayo citado de Elizalde et al. (2003b) un tratamiento adicional con grano entero de maíz generó un aumento de 1,6 kg/día y una eficiencia de conversión de 5,3:1 (Cuadro 7).
El tamaño del grano de maíz obliga al animal a la rumia y lo procesa en la masticación en fracciones menores (quebrado) y fracciones aplastadas (maceradas). Pordomingo et al. (2002a) evaluaron la proporción de grano de maíz en heces en una pérdida del 4 al 9% del grano consumido en una dieta de alto grano y baja fibra. Esas pérdidas pueden ser o no de significa- ción dependiendo de las características del feedlot, experimentalmente no han sido detectadas relevantes sobre el aumento de peso y la eficiencia de conversión. Hibberd et al. (1982), Britton et al. (1986), Reinhardt et al. (1998), Loerch y Fluharty (1998a) y Hejazi et al. (1999), indicaron que, por su tamaño y densidad, el grano de maíz dentado o semi-dentado puede ser utilizado entero sin pérdidas de digestibilidad potencial en dietas de alta concentración de energía metabo- lizable para animales en confinamiento.
Experimentos comparativos de las formas de procesamiento del grano de maíz han en- contrado respuestas similares en aumento de peso al comparar dietas basadas en grano de maíz entero versus molido, partido, aplastado o procesado en copos (Guthrie et al., 1992; Mader et al., 1993; Hill et al., 1996; Mabuku et al., 1996; Loerch y Fluharty, 1998a,b; Reinhardt et al., 1998; Hejazi et al., 1999). En algunos casos se han detectado mejoras en la eficiencia de conversión (Secrist et al., 1996 a, b), y en otros un mayor consumo de materia seca en dietas ofrecidas ad libitum (Guthrie et al., 1992; Bartle y Preston, 1992; Murphy et al., 1994b). Todas las experien- cias coinciden, sin embargo, en que el tipo de maíz utilizado fue harinoso (floury) del tipo dent (dentado y semi-dentado), y de buen tamaño.
Pordomingo et al. (2004) exploraron el uso del grano entero versus molido en dietas de corral con alto grano (60% de maíz, 29% de harina de girasol, 8% de heno de alfalfa y un núcleo vitamínico y mineral) en terneros de destete precoz (60 a 70 días de vida) sin encontrar diferen- cias de significación productiva en tres estratos de peso vivo inicial. Los terneros más pequeños utilizaron con igual o mayor eficiencia el grano que los de mayor peso al destete. Los cuadros 22 y 23 resumen la información de aumento de peso, consumo y eficiencia de conversión de los 123 días de ensayo. Se destaca el alto consumo (relativo al peso) de los terneros en todas las etapas del ensayo.
Comparativamente con las categorías más grandes (novillos), los teneros convierten me- jor el grano entero de maíz. Depetris et al. (2003) reportaron índices de conversión de 4.2:1 (alimento a aumento de peso) para terneras en dietas con dos tipos de grano de maíz entero (alto oleico y tradicional) y 6.4:1 en novillos con la misma dieta (Cuadro 24), similares a las reporta- das por Pordomingo et al. (2004).
En un análisis conjunto de 605 ensayos de alimentación en confinamiento que incluyó información de 22.834 animales, Owens et al. (1997) concluyeron que el potencial del maíz ente- ro para aumento de peso es equivalente al del maíz aplastado en seco o en húmedo, incluso supe- rior al del silaje de grano húmedo, con buenas eficiencias de conversión. Surgió también que la eficiencia energética (estimada como energía metabolizable) del grano de maíz ofrecido entero es superior al ofrecido aplastado. Entre las explicaciones, se argumentó que:
a) el menor contenido de fibra de las dietas de feedlot que incluyen maíz entero, comparadas con las que utilizan maíz aplastado podría inflar el valor del grano entero por transferir al grano una cualidad propia de toda la dieta, menos fibra (Owens et al., 1997);
b) el grano entero promueve una mayor salivación (mayor efecto fibra efectiva) y mayor pH ruminal con lo que se esperaría una reducción de la acidosis subclínica y un mayor consumo (Britton y Stock, 1986; Stock et al., 1995);
c) los efectos asociativos negativos entre el almidón y la fibra en el rumen podrían ser inferiores en dietas con maíz entero que en dietas con grano aplastado o molido, consecuencia de una mayor estabilidad ruminal (Zinn y Owens, 1983); y
d) si la digestión grano no se afecta, el uso de grano entero promueve a un mayor pasaje de partí- culas de almidón sin fermentar en el rumen hacia el tracto inferior con una consecuente mejora en la eficiencia de utilización del almidón (Owens et al., 1986).
Granos húmedos
La utilización de granos húmedos (conservados con humedad en un medio anaeróbico (silos) es frecuente en Argentina. La calidad obtenida, determinada en laboratorio o en ensayos de performance animal, ha sido calificada como similar a la de los granos secos (Clark, 1980; Kennelly et al., 1988; Herrera-Saldana et al., 1990; 1997; Juan et al., 1998; Pordomingo y Juan, 2000).
Estudios de los efectos de las formas de grano húmedo sobre el grado y sitio de digestión han detectado incrementos de la degradabilidad. La conservación con humedad (propia a la co- secha o reconstituida previo al ensilado) es una forma de incrementar la fermentabilidad ruminal de granos con almidón de baja solubilidad como el maíz y el sorgo (McNeill et al., 1976; Galyean, 1976; Hibberd et al., 1982; Britton et al., 1986; Stock et al., 1987ab; Nocek y Tamminga, 1991) y la digestibilidad total, pudiendo ofrecerse enteros, excepto en el caso del sorgo. El aplastado asegura la buena utilización del grano y se recomienda aplastar el grano durante el ensilado.
Juan et al. (1998b) encontraron respuestas similares con grano de sorgo húmedo, grano de maíz húmedo o sus combinaciones en dietas diseñadas para aumento de peso controlado, basadas en 40% de grano y 60% de heno de alfalfa (Cuadros 29 y 30). Debe notarse para los aumentos de peso registrados (1,100 kg/día), la flexibilidad de estos dos recursos en su combina- ción con heno de alfalfa, útil en modelos de alta intensidad en áreas marginales.
Para conservar el grano húmedo sin que se deteriore por putrefacción debemos controlar el desarrollo de microorganismos indeseables (propios de la degradación en presencia de oxí- geno) o fermentaciones a pH superiores a 5. El ensilado es la técnica más difundida. Esta técni- ca aprovecha el desarrollo de acidez por micro-organismos presentes habitualmente en el forraje, cuyo desarrollo se magnifica selectivamente en un medio anaerobio y ácido (el pH común es de 3.8 a 4.5), de fermentación mayoritariamente láctica. Esa fermentación elimina las oxidaciones y fermentaciones de la putrefacción, reduce la tasa de degradación de la materia orgánica y preser- va así el material original en el tiempo.
Debido al alto contenido de almidón del sustrato, un quebrado o aplastado parcial o im- perfecto alcanza para lograr un ensilaje de grano húmedo de buena calidad. Sin embargo, el contenido de humedad es importante para lograr que el grano ensile y se estabilice. El rango óptimo sugerido se encuentra entre 28 y 35% de humedad (agua) en el grano, contenidos cerca del 25% están en el límite inferior por debajo del cual la humedad es escasa para favorecer una rápida fermentación láctica que se extienda, difunda, a toda la masa de grano. Esa carencia de humedad también coincide con compactación escasa y espacios sin grano y con aire en la bolsa o bunker. En el otro extremo, niveles de agua por encima del 35% no se retienen en la masa del grano y ese excedente de agua tiende a bajar hacia el piso de la bolsa o bunker, lavando sustan- cias ácidas y nutrientes. Entre ambos extremos, el segundo tiene mejores chances de preservar el grano que el primero.
Cuando la generación de ácido láctico es lenta y escasa y la remoción del oxigeno es parcial por falta de compactación, ausencia de almidón o carbohidratos rápidamente fermenteci- bles, la acidez tarda en desarrollar y el pH en decrecer por debajo de 5, aumentan las fermenta- ciones butílicas y oxidaciones parciales, el material se mantiene a mayor temperatura debido a esas oxidaciones, cambia de color (torna hacia formas más oscuras), con degradación de azúcares y proteínas y se deteriora progresivamente la calidad. Ese tipo de silajes es menos estable en calidad y palatabilidad y puede limitar el consumo. Si se desarrollan sectores de putrefacción se pueden generar toxinas de orgen fúngico (aflatoxinas, trichotezenes, etc.)
Una técnica alternativa menos utilizada es la conservación en un medio alcalino. Esta opción no requiere de la anaerobiosis necesaria en el silaje y el cambio de color del grano. La masa de grano se puede alcalinizar mediante el agregado de urea (2 al 3 % de la materia seca de grano a conservar) durante la descarga del grano de la tolva o camión que lo transporta recién cosechado. Para esta forma de conservación el contenido de humedad deseable se debería ubicar entre el 30 y el 35% (65 a 70% de materia seca). Se almacena el grano a granel bajo un tinglado, en galpón, o en un silo aéreo con malla metálica o plástica para granos. Con la humedad propia del grano se solubiliza la urea, hidroliza y convierte en amoníaco, el cual da origen a hidróxido de amonio, elevándose el pH 8 o más.
Ese medio alcalino impide el desarrollo de la microflora fúngica y bacteriana responsa- ble de la putrefacción (Ghate y Bilansky, 1981; Russell et al., 1988; Russell y Schmidt, 1993). Es importante que el contenido de humedad no sea inferior al 28%, particularmente en sorgos (Juan et al., 1998a; Pordomingo y Juan, 2000). A niveles de 25% de humedad o inferiores, la difusión del amoníaco es despareja, la alcalinidad lograda no alcanza a pH 8 y la acción sobre el pericarpio del grano es limitada. En ese estado no se eliminan los hongos y en pocos días apare- cen sectores de putrefacción que rápidamente toman a la totalidad de la masa de grano (Pordo- mingo et al., 2002c). Si la disolución de la urea e hidrólisis en amoníaco ha sido adecuada, a los dos o tres días de aplicada la urea, el grano debería mostrar una apariencia húmeda y brillante, untuosa al tacto, que tiñe o mancha con pigmentos del pericarpio (Juan et al., 1998a; Pordomingo y Juan, 2000; Pordomingo et al., 2002c).
En contraposición al ensilado anaeróbico, al no ser exigente en anaerobiosis, esta técnica implicaría un ahorro en la estructura de conservación y facilitaría la tarea de producción de la reserva. Existen antecedentes que indican que el amoníaco también provocaría un ablandamien- to del pericarpio del grano, lo que haría innecesaria la molienda o partido del grano para lograr una buena digestión en el bovino (Russell y Schmidt, 1993).
En dietas con 78% de grano de maíz conservado con urea, con grano húmedo ensilado o grano seco (Cuadro 11), Pordomingo et al. (2002b) reportaron consumos, aumentos y eficiencias de conversión similares entre tratamientos para las formas húmedas comparada con la forma seca y molida tanto en vaquillonas como en novillos (Cuadro 12, 13 y 14). En particular, el trata- miento que incluyó el maíz húmedo conservado con urea superó a los otros dos en aumento de peso y eficiencia de conversión (Cuadro 15). El agregado de urea enriqueció de nitrógeno al grano de maíz (Cuadro 11) y a la dieta por lo que no se agregó urea adicional para completar los requerimientos de proteína bruta (Cuadro 11). En los Cuadros 16 y 17 puede observarse la com- posición nutritiva de los insumos utilizados y la evolución del contenido de materia seca y del pH del grano conservado con urea. Esta forma de conservación resulta menos estable que la forma de ensilado ácido en anaerobiosis y la preservación por períodos prolongados sería una limitante de esta técnica de conservación.
Aunque el nivel de consumo durante el acostumbramiento y los primeros 50 días resultó superior para los animales con la dieta de grano seco y molido, comparado con el consumo de los tratamientos con granos húmedos (Cuadro 15), los animales se acostumbraron a las dietas húme- das y el menor consumo inicial no se reflejó en menor ritmo de engorde. Por el contrario, dicho efecto fue el responsable de la mejor eficiencia de conversión registrada para las formas húmedas y en especial para el tratamiento que incluyó grano conservado con urea. El olor a amoníaco, los cambios de color (oscurecimiento) y la aparente astringencia de la untuosidad del grano no pare- cieron afectar el nivel de consumo luego de acostumbrados los animales a la dieta, comparado con el consumo de los animales con grano conservado húmedo ensilado sin urea. La conserva- ción en medio húmedo y alcalino (amoníaco generado a partir de la hidrólisis de la urea) permi- tió también lograr una alta digestión del grano haciendo innecesario el procesado del mismo cuando se lo ofrece en dietas de alto carga de almidón (78% grano).
En sorgo conservado con urea se encontraron respuestas similares a las observadas en maíz (Juan et al., 1998a; Pordomingo y Juan, 2000), siempre que el grano fuese aplastado previo a ser ofrecido o conservado con quebrado previo (Pordomingo y Juan, 2000). De lo contrario, la pérdida por grano entero no digerido es muy elevada. Estos autores encontraron que para los granos húmedos de sorgo con taninos es insuficiente el efecto erosivo del pericarpio del grano que produce el amoníaco para degradar la cubierta y exponer el almidón a los procesos digesti- vos (Pordomingo et al., 2003). El grano entero de sorgo resultó en el menor aumento de peso y la peor eficiencia de conversión.
Más de un tipo de grano
La mezcla de granos genera mejor conversión de alimento a aumento de peso (Fulton et al., 1979; Kreikemeier et al., 1987; Stock et al., 1987a,b; Gross et al., 1988). Pordomingo et al. (2002b) reportaron mejor aumento y conversión cuando se agregó grano entero de avena a una dieta basada en grano de maíz. El Cuadro 18 resume la información de la composición de las dietas utilizadas en ese ensayo. En el Cuadro 19 se muestra la composición nutritiva de los in- gredientes utilizados y el Cuadro 20 los resultados de producción y eficiencia.
Las tendencias observadas en T3 (Cuadro 20) sugieren que la mezcla de granos de maíz y avena en proporción 80:20 permitiría lograr una mayor performance que cuando se usa grano de maíz solamente. Ello podría justificarse por: a) un mejor balance de calidad de almidones, b) mayor estabilidad en la tasa de fermentación y digestión del alimento, c) el mayor contenido de proteína en el grano de avena y c) la alta fermentabilidad ruminal del grano de avena. Beneficios de las mezclas de almidón de distinto origen han sido reportados por Huck et al. (1998), Kreikemeier et al. (1987), Bock et sl., (1991) y Streeter et al. (1989). Los motivos se centran en los efectos de la mezcla de almidones de diferente tasa de degradabilidad en el rumen y fracciones pasantes hacia el tracto inferior, la complementariedad de la composición de la fracción proteica de los granos, y los efectos sobre la palatabilidad y textura de la dieta.
El almidón del grano de sorgo es el de menor tasa de degradación en el rumen y el de mayor escape a la fermentación ruminal, seguido por el maíz (Hibberd et al., 1982) y luego por los otros granos (avena, cebada, trigo). Estos últimos contienen almidones de mayor degradabi- lidad ruminal que los de verano (Morgan y Campling, 1978; Phillippeau et al., 1999b). Sin embargo, la variedad o híbrido, en tamaño y el tipo de grano (ej. harinoso o dent y córneo o flint en el maíz), tienen influencia en la degradabilidad ruminal del almidón (Philippeau et al., 1999b; Maresca et al., 2002). La mezcla de almidones de distinta tasa de degradabilidad genera un am- biente ruminal de fermentación más estable, y la fracción de escape permite una absorción de una fuente energética en intestino cuyo rendimiento energético es superior al logrado si fuese totalmente fermentado en rumen.
Por su parte la complementariedad de las fuentes proteicas de los granos aporta un mejor balance de aminoácidos y péptidos en el rumen reduce la dependencia de la síntesis de aminoáci- dos esenciales en el rumen y mejora la tasa de crecimiento de las poblaciones microbianas (Ken- nelly et al., 1988; Huck et al., 1998).
La mezcla de texturas de granos en la dieta reduce la presencia de polvos, mejora la toma (aprehensión) por el animal y reduce el rechazo de los granos menos palatables (Huck et al., 1998; Kreikemeier et al., 1987) especialmente cuando se mezclan granos molidos con enteros (Mabuku et al., 1996; Harmon et al., 1987; Lee et al., 1982) (ej. sorgo molido con avena entera; maíz aplastado con avena entera, sorgo molido con maíz aplastado; grano húmedo de maíz ente- ro con maíz molido o sorgo molido; grano húmedo de maíz con avena entera; maíz aplastado con avena aplastada; grano molido de sorgo con trigo entero, granos de maíz aplastado con ceba- da aplastada; cebada entera y sorgo molido). Entre los granos finos, la cebada aplastada es el que mejor se complementa con las formas procesadas del maíz y del sorgo (molidos, quebrados o aplastados).
La fibra
La fibra en dietas de corral de alto contenido de almidón ejerce un efecto físico o mecá- nico más que nutritivo. El valor alimenticio de la celulosa (fibra) en esas dietas es muy bajo (Swingle, 1995), principalmente por la baja degradación ruminal que ocurre con esa fracción en dietas de alta proporción de concentrado (Zinn y Owens, 1983). El ambiente ruminal con alta carga de almidón es demasiado ácido (pH = 5,0 a 5,5) para el desarrollo de bacterias celulolíticas en cantidad suficiente para digerir eficientemente la fibra (Pordomingo et al., 1999b). El princi- pal objeto de la fibra en estos casos es el de reducir la tasa de consumo y promover la rumia, la salivación y la consecuente producción de buffer ruminal para disminuir el riesgo de acidosis (Kreikemeier et al., 1990).
Dependiendo del contenido de fracciones fibrosas en los otros componentes de la dieta, la cantidad mínima del recurso fibroso a incorporar en se ubica comúnmente entre el 5 y el 10% de la dieta (base seca). Para sostener una actividad fermentativa adecuada se sugiere un aporte tal de elementos fibrosos que aseguren un mínimo de 10% de fibra detergente ácido (FDA), y al menos la mitad de ese aporte provenga de una fuente de “fibra efectiva” o larga tal como los henos o los silajes. Sin embargo, el efecto de “fibra efectiva” deseado no radica en el aporte energético de esa fracción sino en el efecto mecánico por lo que también se usan sustitutos de menor costo que el heno que cumplen funciones similares como las cáscaras de semillas y resi- duos fibrosos de la industria de los alimentos.
El oferente de fibra más común utilizado en la mayoría de los sistemas alimentación en confinamiento es el heno (rollo o fardos) procesados picados o molidos gruesos y de calidad intermedia o baja. La calidad y tipo de fibra es de escasa relevancia en las dietas de alto grano pero existen recursos fibrosos que se adaptan mejor que otros, dependiendo del tipo de grano en que se basa la dieta (Theurer et al., 1999).
Otros oferentes de fibra son los silajes de planta entera (ej. Silaje de maíz, silaje de pas- turas o alfalfa, silajes de ryegrass, avena o cebada). Algunos de éstos son también buenos oferentes de energía por su contenido de grano (silajes de maíz, sorgo, avena o cebada) (Petit y Tremblay, 1995; Castro et al., 2002; Pordomingo, 2002; Santini et al., 2003) o la alta digestibili- dad total (silaje de ryegrass) (Cardoniga y Satter, 1993; Zaman et al., 2002). En estos casos, la proporción incorporada es mayor que la del heno porque se pretende un aporte energético y no solo mecánico.
Aunque la cantidad de fibra necesaria en los planteos de feedlot es baja, su provisión es una limitante operativa y económica. El heno cosechado en rollos y luego procesado para su mezclado en la dieta, se convierte en uno de los insumos más caros por unidad de energía diges- tible. Incluso, en la mayoría de los feedlots de poca escala, no se dispone de equipamiento para molienda de henos. Sostener una fermentación adecuada con independencia de fuentes de fibra larga, permite simplificar la alimentación a corral y hacer accesible esta práctica a numerosos planteos de engorde. En este extremo, sin embargo, los riesgos de acidosis son mayores.
En un nivel intermedio entre los planteos con heno como fuente de fibra y los sin fibra larga (ver más abajo), se encuentran aquellos que incorporan fibra a través del afrechillo de trigo, la raicilla de malta y las cáscaras (cáscara de girasol, de semilla de algodón o maní). De todas las cáscaras, las menos “leñosas” o duras serán las mejores desde el punto de vista de su fermenta- ción y tolerancia a través del tracto digestivo. En proporción de 10 a 12% de la dieta, la cáscara de maní es muy utilizada para reemplazar totalmente la fuente de fibra larga (henos). Ofrecida sin moler, se le atribuye alta efectividad en generar el efecto fibra para mantener la motilidad ruminal (Utley et al., 1973). Las otras cáscaras incrementan volumen pero son menos efectivas en generar el efecto fibra. La cáscara de girasoles grandes, del tipo rayado, tienen mayor efecto que la de tipo negro para aceite, más pequeñas. La cáscara de semilla de algodón o de soja no aportan al efecto fibra por lo que no pueden reemplazar a la fuende de fibra larga totalmente.
La cáscara de soja es de mayor calidad que las otras, tiene alto contenido proteico y su digestibilidad es muy superior a la de las de girasol o maní (Cuadro 2). Se la suguere para reem- plazar el maíz aplastado en recría en proporciones del 30 al 60% sin resentir el aumento de peso (Mueller et al., 2011). El afrechillo de trigo (en pellet) y la raicilla de malta pueden utilizarse para diluir la energía metabolzable de las dietas y generar efectos similares a los de la fibra, pero tales efectos son factibles a niveles de inclusión alto (superior al 20%) y debe tenerse en cuenta que estos son también oferentes de proteína bruta (Elizalde et al., 2003a). Tienen además un buen nivel energético y buena digestibilidad.
La inclusión de cáscaras, especialmente la de maní, requiere de conocer la composición la misma para lo cual es necesario recurrir al análisis de laboratorio. Siendo una fracción fibrosa del fruto, se espera que tenga un alto contenido de fibra (FDN y FDA) y un contenido bajo de proteina, pero los datos más relevantes son el contenido de materia seca (% MS) y el de cenizas. El primero tiene que ver con la cantidad de agua que tiene la cáscara. Este subproducto debería ser muy seco (más de 88 % de MS). Si tiene agua, puede haber ocurrido el agregado para “aplas- tar” y compactar la carga y transportar más peso, pero por el contenido inicial de tierra (fruto hipógeo) la humectación de la cáscara promueve el desarrollo de hongos y sus toxinas (aflatoxi- nas, trichotecenes, etc). Si el valor de cenizas es muy elevado (superior al 6%) podría presupo- nerse un enriquecimiento con tierra durante la carga o el barrido de la cáscara (por encima del esperable de una cáscara que inicialmente es cosechada mezclada con suelo). La cáscara limpia no debería superar el 3 % de cenizas.
Existen evidencias experimentales (Pordomingo et al., 2002a; Elizalde et al., 2002; Ma- resca et al., 2003) y de productores en Argentina que coinciden en que es posible alimentar a corral con dietas sin fuente de fibra (sin heno o silaje), basadas solamente en grano, harina de girasol o de soja y un complemento mineral y vitamínico con un ionóforo (monensina o lasalo- cid).
Murphy et al. (1994a) no encontraron diferencias en la digestión de almidón proveniente de una dieta basada en maíz entero y sin fibra, comparada con una equivalente con maíz aplasta- do, cuando la oferta de alimento no fue restringida. Resultados que fueron confirmados en Ar- gentina (Pordomingo et al., 2002b; Maresca et al., 2002). Sin embargo, cuando se restringió la oferta de alimento al 70% del consumo voluntario, la digestibilidad del almidón proveniente del grano entero fue inferior a la del almidón de grano aplastado. De manera similar, Turgeon et al. (1988) encontraron diferencias en la digestibilidad del grano de maíz cuando alimentaron novi- llos con maíz entero o partido en dietas sin fibra larga y a 2,3 veces el nivel de mantenimiento.
Pordomingo et al. (2002b) compararon dietas con grano entero de maíz y harina de gira- sol o grano entero de maíz y avena, y harina de girasol, sin fuente fibra larga, versus dietas con la inclusión de 10% de heno de alfalfa (Cuadros 18, 19 y 20). Los planteos sin fibra igualaron (y tendieron a superar) en aumento de peso al tratamiento con heno. No se detectaron efectos aci- dóticos o trastornos visibles en los tratamientos sin fibra.
Entre las alternativas en el suministro de fibra en la forma de heno, se encuentran las cáscaras de semillas (cáscara de semilla de algodón, de semilla de girasol, de soja, de arroz o de maní), residuos fibrosos de la agro-industria (raicillas, afrechillos, etc.) Aunque existen asocia- ciones entre tipos y procesado del grano, y el tipo de fibra a utilizar (Owens et al., 1997). Las fuentes de fibra antes citadas son utilizables sin diferenciarse en el resultado sobre aumento de peso o eficiencia de conversión, pero el efecto de fibra efectiva es mayor en las cáscaras de ma- yor tamaño. La cáscara de maní es más efectiva que la de girasol. Dentro de la variedad de cás- caras de girasol, las del tipo “confitero” (grande) tienen mayor efectividad que las de girasol para industria. Las cáscaras de girasol para industria, de soja y de arroz tienen un efecto marginal y limitado para sustituir a otras fuentes de fibra efectiva.
Raciones a base de cebada (75 a 85%) entera resultan en similares resultados productivos al maíz (Elizalde et al., 2003b). La inclusión de cebadas con alta proteína permite reducir la necesidad de fuentes proteicas adicionales y, por su contenido de fibra, el requerimiento de fibra extra es también menor. Ofrecida entera podría conformar raciones completas con un 10% de harina de girasol, y un núcleo vitamínico y mineral al 2 o 3% (depende del núcleo). Aunque el aporte proteico de grano es variable (9 a 15% de proteína bruta), con un contenido medio de proteína bruta de 12% y un 30% en la harina de girasol (pellet) se logra una ración con 14,5% de proteína de alta metabolicidad que cubre los requerimientos de la mayoría de las categorías.
Pordomingo et al. (2007a) evaluaron una dieta basada en 96% cebada sin una fuente de fibra adicional lográndose aumentos y conversiones similares a las obtenidas con una dieta basa- das en cebada y afrechillo de trigo. El oferente de nitrógeno (urea o afrechillo de trigo) generó diferencias a favor a la dieta que incluyera la fuente de proteina verdadera (Cuadro 52). Durante la primera etapa (50 días) la dieta de cebada fue ofrecida entera y durante la segunda (40 días) fue ofrecida aplastada. Estas observaciones, coincidentes con otras (Koenig y Beauchemin, 2013a), indicarían que la cebada es un grano de buena versatilidad para reemplazar a otros gra- nos, y reducir la demanda de concentrados proteicos y de fibra.
En una experiencia con novillos de 253 kg al inicio y más de 400 kg a la terminación, Pordomingo et al. (2011) reportaron 100 g mas de aumentos de peso diario con dietas basadas en maíz y cebaba aplastados, sin fuente de fibra, ni concentrado proteico, o con mínimo aporte del mismo (6% de harina de girasol) (Cuadro 53, versus una dieta basada en grano de maíz (77%), harina de girasol (12%) y afrechillo de trigo (8%). Se destaca en esta experiencia la con- versión de las dietas, la complementariedad de los granos y el efecto en reducción de oferta de proteina verdadera que permite la inclusión del grano de cebada en proporciones importantes en la dieta.
Elizalde et al. (2003a) estudiaron la inclusión creciente de afrechillo de trigo en dietas a base de cebada aplastada y lograron aumentos de peso vivo (1,500 kg/día) y conversiones simila- res (6:1) con 75, 65 o 55% de cebada aplastada y cantidades crecientes de afrechillo (12,5, 35 o 45%) (Cuadro 25). La combinación de la cebada con el afrechillo de trigo permitió mantener constante el consumo de energía metabolizable y la eficiencia fermentativa. La tasa de engrasa- miento y el engrasamiento final en ese ensayo resultó también similar entre tratamientos (Cuadro 15). Se destaca la calidad del afrechillo de trigo para complementar dietas de alta energía.
Pordomingo et al. (2007b) incluyeron 22% de afrechillo de trigo en una dieta basada en grano de maiz, reemplazando la fuente de fibra larga (heno de alfalfa) y la mitad del oferente proteico (harina de girasol) (Cuadro 54), ofrecida a terneros en confinamiento. Ambas dietas fueron equivalentes en proteína bruta, fibra detergente ácido y oferta de energía metabolizable. El resultado en aumento de peso (1,200 kg/día) y el consumo relativo al peso vivo medio (2.6%) fue similar entre ambas dietas (Cuadro 54).
Silajes
El silaje de planta entera de maíz o de sorgo se incorpora a dietas de engorde a corral. Su participación en las dietas depende de las metas en cuando a aumento de peso. El silaje no es un oferente de energía del nivel del grano, su concentración de energía metabolizable se asemeja a la que puede proveer un verdeo de invierno o una pastura en primavera. Por ello, los encierres basados en silajes de planta entera (ej. 80% silaje de maíz + 20% harina de girasol) permiten aumentos de peso de 700 a 1000 g/día, pero difícilmente mayores (Santini et al., 2003; Castro et al., 2002; Juan et al., 1997). Estos aumentos son útiles para planteos que pretenden encerrar sobre la base de un alto rendimiento de materia seca y alto aprovechamiento de la misma, pero que no tienen como meta muy altos ritmos de engorde, terminaciones de alto engrasamiento o muy rápidas.
Existen combinaciones intermedias enriquecidas con grano, donde el 40 a 50% de la die- ta es silaje de planta y el resto se completa con grano (20 a 30%) y la harina de girasol o soja (15 a 25%). Estas dietas son energéticamente algo superiores a la de silaje puro, pero todavía des- cansan en la fermentación de una buena cantidad de fibra proveniente del silaje. El aumento puede ser algo superior al antes citado, pero no alcanza los niveles de un engorde típico basado en grano. Son, sin embargo, las sugeridas en recría con aumento controlado, encierres de acopio de terneros, encierres de post-destete, para planteos que luego terminan a campo en pastoreo o de vacas en engorde.
Santini et al. (2003) reportaron aumentos de peso similares entre dietas con 78% silaje o 33% de silaje y 45% de maíz húmedo en novillos británicos de biotipo grande (1,07 kg/día) o de biotipo chico (0,89 kg/día) (Cuadro 26). Tampoco detectaron diferencias dentro del biotipo grande en tasa de engrasamiento y engrasamiento final. En el biotipo chico el espesor de grasa dorsal fue superior para la dieta con más grano y también la tasa de engrasamiento (Cuadro 25).
Castro et al. (2002) detectaron mejoras en el ritmo de engorde con el incremento de grano en la dieta, pero no demasiado importantes en términos absolutos (1,129, 1,252 y 1,325 kg/día para alto, medio y bajo contenido de silaje) en novillos en terminación (Cuadro 27). Estos resultados dependen del contenido energético del silaje de maíz, pero expresan su plasticidad y potencial.
La calidad del silaje puede fluctuar entre años y lotes de manera significativa. Para poder predecir la performance de los animales es necesario contar entonces con información de la cali- dad nutritiva. Santini y Paván (2002) compararon resultado de aumento de peso entre años en novillitos alimentados con silaje y harina de girasol, y encontraron que el aumento de peso puede variar en un rango de menos de 800 gramos/día a casi 1 kg, dependiendo particularmente de la oferta de almidón del silaje (Cuadro 28).
El contenido de humedad y de grano y el momento de picado son muy relevantes en la calidad final del silaje de maíz o de sorgo. Los silajes con mucho agua (más del 75%) o, lo que es lo mismo, bajo contenido de materia seca (inferior al 25%) son silajes que limitan el consumo y generan mucho efluente líquido, con la consecuente pérdida de nutrientes (carbohidratos solu- bles y proteínas), se lava también el ácido láctico y, aunque su picado es generalmente bueno, puede limitarse la calidad de la fermentación por exceso de agua. En el otro extremo, silajes con más de 40% de materia seca al momento de la confección (picado y ensilado), tienen mayor can- tidad de grano, pero se pican con dificultad, queda un picado muy heterogéneo y compactan mal, quedando demasiado aire en la masa de silaje.
El exceso de aire promueve las oxidaciones y las fermentaciones butíricas, la temperatu- ra permanece alta por respiración y el pH no baja de 5. Ese material se deteriora con el transcur- so de los días porque no se logra eliminar la respiración de los carbohidratos (almidón) y alcan- zar concentraciones de ácido acético y láctico que estabilice el ensilado. Entre los cambios más visibles, se observa el desarrollo de coloraciones oscuras (marrones y negros), desintegración de la estructura original del material (se torna dificultoso diferenciar las fracciones de planta) y el olor es característico de fermentaciones butíricas, mezclado con óxidos de azufre. Se puede ob- servar humedad y formación de gotas en el material, consecuencia de la oxidación y producción de agua por acción fúngica y bacteriana. Este tipo de silaje debería ser evitado en dietas con más de 50% de silaje (base seca), en animales de tambo y en terneros que inician el engorde, pero puede participar en dietas de alto grano y fracciones moderadas de silaje (menos de 20% de silaje en base seca). Debe verificarse que no se afecte la palatabilidad total o aparezca micelio de hon- gos (masa fúngica) muy difundido.
El contenido de grano mejora la oferta de energía metabolizable y es clave en la perfor- mance animal en dietas a base de silaje. En los silajes con poco grano (menos del 30% en base seca) la calidad depende de la digestibilidad de la fibra (tallo y chala del choclo). Para mejorar el contenido de grano en cultivos con mucho follaje o de poco rendimiento podría elevarse la plata- forma de recolección hasta el nivel de la base de las espigas y dejar parte de tallo sin cortar. Esto permite elevar la relación grano/planta picada en un 20 a 30%, por lo que un cultivo para silaje podría pasar de 25% a 35% de grano en el material cortado y cambiar la digestibilidad y la con- centración energética del silaje.
La inclusión de grano a silajes de planta enera es una herramienta para mejorar la oferta de energía metabolizable de la dieta basada en silaje. La mejora de oferta energética con el in- cremento de la proporción de grano es muy significativa cuando los silajes son de forrajeras sin grano (pasturas, alfalfa, verdeos cortados en estado vegetativo). Las dietas en base a silaje de alfalfa son las que mejor se complementan con la adición de grano pasando de una concentración energética limiada, para mantenimiento o aumentos de peso menores a los 500 g/día en la mayo- ría de los casos, a una oferta de energía compatible con engordes de terminación (800 a 1000 g/día). En este escenario, el grano se incorpora reeemplazando casi la 1/3 de la dieta.
Varias evaluaciones han mostrado los efectos positivos del enriquecimiento con grano de las dietas basadas en silajes, pero también han advertido que el incremento del potencial o la mejora de la respuesta individual (aumento de peso) no es lineal. En dietas basadas en silaje de maíz, Pordomingo et al. (2012) demostraron que el la inclusión de grano molido de maíz mejora la respuesta individual hasta el punto en que el grano en la dieta (ofrecido por el mismo silaje más el agregado) contribuye el 36% de la dieta (en base seca) (Cuadro 56). Enriquecimiento con una mayor cantidad de grano no generaría respuestas mejores en aumento de peso, hasta que el grano contribuye la mayor fracción de la dieta.
Se hipotetiza que el incremento creciente de grano aumenta la digestibilidad total de la dieta, pero el cambio de la fermentación ruminal hacia una de naturaleza amilolítica deteriora o retarda la fermentación de la fibra en el rumen. Como pas dietas con alta proporción de silaje requieren de fermentación de la fibra para su utilización eficiente, la mejora de la oferta de ener- gía metabolizable con el agregado de grano, encuentra en contraposición un deterioro de la oferta energética por la degradación de la fibra.
En los casos en los que el aporte de grano total constituye a esta fracción en el ingredien- te mayoritario, la dieta se asemeja a una dieta de alto grano y baja fibra e importa menos la ener- gía a obtener por la fermetnación de la fibra proveniente del silaje. La fibra pasaría tener un rol físico o mecánico en el proceso de digestión, y de menor relevancia como nutriente. De todas formas, la calidad y buena conservación del silaje son igualmente relevantes por sus impliancias en la palatabilidad y el consumo.
En el ensayo reportado de Pordomingo et al. (2012) puede observarse el potencial de en- gorde del silaje de maíz. Aún a contenido bajo de grano (24% de la materia seca total de la die- ta), la dieta con 85% de silaje de maíz, correjida por proteina y minerales, alcanzó a un engode de 750 g/día en novillos en terminación. Este aumento de peso se incrementó a 1000 g/día cuan- do se incrementó la proporción de grano al 36% (Cuadro 55). En otra experiencia basada en sila- je de planta entera de sorgo granífero con 62.5, 70 0 77.5% de silaje (en base seca), Volpi La- greca et al. (2010) mostraron el pontencial de los silajes para el engorde de novillos. Aun la dieta con más silaje y urea como oferente adicional de nitrógeno se lograron aumentos de peso de 992 g/día, y terminación de novillos a 418 kg de peso vivo (Cuadro 56). El consumo llegó a 2.56% del peso vivo para ese tratamiento. Ls oferta de grano en ese silaje era del 39% (base seca).
En las dietas típicas de corral de alto grano y baja fibra el silaje de planta entera es utili- zado para reemplazar la fracción de heno (u otra fuente de fibra). Participa en proporciones del 10 al 20% del alimento (en base seca). En estos casos, su inclusión sirve también para aportar humedad a la dieta, sobre la que se adhieren mejor los otros componentes, también aumenta el volumen y mejora la homogeneidad del consumo. El silaje es más atractivo que el heno de baja calidad y hace un aporte adicional como “buffer” (amortiguador) del pH ruminal (Owens et al., 1997). Sin embargo, debido a la baja proporción en que se incluye el silaje en estas dietas, la incorporación al sistema se justifica si el tamaño del feedlot hace factible la infraestructura adi- cional que impone la confección, almacenado y extracción del silaje.
Entre otros silajes probados en Argentina debemos incluir a los de sorgo granífero y los de avena con grano. La relación grano planta es más elevada en estos que en el silaje de maíz. La digestibilidad del silaje de sorgo puede resultar inferior a la del silaje de maíz, por menor digestibilidad del grano y de la planta de sorgo, pero la mayor proporción de grano en el silaje de sorgo puede compensar la menor digestibilidad promedio. El silaje de avena (confeccionada en estado pastoso del grano) provee es similar al del silaje maíz. El rendimiento por hectárea, sin embargo, es menor en este recurso.
Asimismo, la combinación de silaje de maíz y heno de calidad, complementados con grano de maíz, es una alternativa muy flexible para lograr aumentos de peso estables en planteos de aumento controlado. Juan et al. (1997) reportaron los resultados de un estudio de dietas hú- medas (a base de silaje) o secas (a base de heno de alfalfa), y sus combinaciones con adición de grano y harina de soja. Las dietas fueron equivalentes en oferta de proteína bruta y energía me- tabolizable y generaron aumentos de peso similares en vaquillonas alimentadas a corral (Cuadro 31). Parra et al. (2002) reportaron similares aumentos de peso para dieta con menos de 60% de grano a base de silaje (Cuadro 32). En estos planteos con altos niveles de fibra larga, sería con- veniente procesar el grano de maíz para mejorar la utilización del almidón.
Maresca et al. (2003) probaron que con el incremento del nivel de fibra se reduce la utili- zación del almidón del grano entero de maíz, efecto que se magnifica con la edad de los animales (Cuadros 36 y 37). Puede observarse en el Cuadro 37 que la digestibilidad del almidón es mayor en el tratamiento con baja fibra y que en ambos tratamientos empeora con el aumento de la edad del animal. La utilización de la fibra de la dieta también decrece con la edad y es inferior en el planteo con alto contenido de grano (resultado esperable por la mayor incidencia de la fermenta- ción amilolítica, depresora de la fibrólisis).
El silaje de gramíneas o leguminosas tiene un potencial energético menor que el que maíz o sorgo, pero puede ser de utilidad en planteos de recría. En planteos de novillo pesado, Steen et al. (2000) reportaron aumentos de peso de 700 y 800 gramos/día cuando se combinó el silaje de ryegrass con avena aplastada (Cuadro 32). En animales más livianos sería esperable un aumento mayor y una conversión más eficiente de estos recursos.
Por su parte, trabajos en Argentina de alimentación a corral con dietas basadas en silajes de sorgo del tipo silero o forrajero (bajo contenido o nada de grano) indicarían que, dependiendo de la calidad de la fibra y el contenido de carbohidratos solubles, estos materiales tienen poten- cial para generar engordes de 800 g/día o superiores. Kent et al., 2011) reportaron resultados de una comparación de engorde novillos sobre silajes de sorgo granífero, silero y forrajero. Vaqui- llonas y novillos sobre dietas basadas en sorgo grasnífero tuvieron los aumentos de peso mayores (928 y 2089 g/día, respectivametne), pero los animales sobre sorgo forrajero tuvieron aumentos de 806 y 991 g/día (vaquillonas y novillos, respectivamente). Estas respuestas fueron considera- das muy buenas para forrajes que pueden producirse en ambientes restrictivos de la producción de granos o silajes graníferos (Cuadro 57).
El concentrado proteico
El oferente proteico participa en las dietas de feedlot en el mínimo necesario para lograr el aporte de proteína que la categoría animal requiere. En el cálculo debe tenerse en cuenta el contenido de proteína de cada insumo, incluido el del grano, el que aunque bajo si se trata de maíz o sirgo (7 a 9%), es importante por la fracción mayoritaria que ocupa de toda la dieta. Los concentrados proteicos más comunes en la región pampeana son las harinas de girasol y de soja. La semilla de algodón y el gluten feed (sub-producto proteico de la industria del maíz) son tam- bién utilizados en las regiones del centro y norte del país. Las harinas proteicas citadas difieren en la cantidad de proteína bruta que aportan (ej. la harina de girasol ofrece de 28 a 32% y la de soja de 42 a 48%) por lo que sería conveniente tener información de laboratorio de la proteína bruta contenida en los insumos que pensamos usar.
Una alternativa son los concentrados proteicos comerciales, los que por su alto contenido de proteína bruta (generalmente por encima del 40%), permiten reducir la cantidad de este insu- mo a un mínimo y permiten e incrementar la cantidad de grano al máximo tolerable. Es conve- niente que estos concentrados contengan el complejo vitamínico y mineral, incluyendo un ionó- foro (entre los más comunes: monensina y lasalocid) que opera de modulador de la fermentación y del consumo (ver más adelante).
En el Cuadro 1 se puede observar que la proporción de harina proteica es menor en los casos donde se incluyó pellet de soja, comparados con los casos donde se incluyó harina (pellet o harina) de girasol. Sin embargo, la proporción de proteína bruta fue similar. La harina de soja tiene más proteína bruta que la harina de girasol. Entre otros detalles puede resaltarse que el contenido de FDA (fibra) es similar en todas y próximo al 10% (valor mínimo deseable) del ali- mento (base seca), del cual al menos el 60% es fibra efectiva proveniente del heno (rollo o fardo molido o desmenuzado
Cuando se incluye un grano de invierno como la avena, disminuye también la necesidad de harinas proteicas porque el grano de avena hace un aporte de proteína mayor que el del maíz. Puede observarse que la participación de la urea permite una reducción importante del uso de estas harinas (pellets) (dietas T1a T4 versus T5), pero en ningún caso supera el 1% de la formu- lación total. Niveles de 1 a 1,2% de urea son considerados el límite superior de inclusión de urea en dietas de feedlot sin riesgo de intoxicación amoniacal. Puede observarse también que todas las dietas excepto T2 han sido formuladas incluyendo un macro núcleo vitamínico y mineral. En T2 incorpora un micro núcleo y se aportan los macro minerales (calcio, magnesio, fósforo y la sal) por separado.
Si se evalúan estas dietas desde su concentración energética u oferta de energía metabo- lizable son dietas para aumentos diarios de peso vivo de 1,300 a 1,500 kg, pero la oferta de pro- teína metabolizable puede resultar limitante de ese aumento y el componente específico en défi- cit será la fracción de proteína no degradable en rumen (proteína pasante) (Ainslie et al., 1993; Knaus et al., 1998; Stock et al., 1989; Titgemeyer et al., 1989).
En los sistemas actuales es difícil en términos económicamente factibles incorporar pro- teínas de baja degradabilidad ruminal. El recurso más común en el pasado lo fue la harina de carne. Hoy el uso de harina de carne, de hueso u otras de origen bovino está prohibido debido al riesgo de transferencia de enfermedades como BSE (vaca loca). Otros recursos son las proteínas vegetales tratadas con calor o agentes químicos (Spears et al., 1980) para reducir su solubilidad y en consecuencia su fermentación ruminal, tales como harina de soja tostada, soja entera tostada, harina de soja tratada con formaldehído (Stern et al., 1994). Pero, estos recursos son caros y de escasa relevancia en el feedlot argentino (utilizados en feedlots de Norte América, Australia, o Europa.)
Sin embargo, la principal limitante de la eficiencia de utilización de la proteína es la me- tabolicidad de la dieta y el balance de proteína metabolizable (Meissner, 1992; Ainslie et al., 1993; Castillo et al., 2001a,b). En dietas basadas en proteínas degradables en rumen, el balance de proteína metabolizable puede ser neutro e incluso positivo, por lo que el efecto adicional so- bre la eficiencia de conversión con el suministro de proteínas de baja degradabilidad ruminal es bajo o nulo (Loerch y Fluharty, 1980; Beever, 1993). La sincronía de la fermentación ruminal entre almidón y proteína es el mayor contribuyente a la metabolicidad de la dieta (Herrera- Saldana et al., 1990 b; Hoover y Stokes, 1993) y eficiencia de la síntesis y provisión de proteína microbiana al tracto inferior (Sinclair et al., 1995; Koenig y Beauchemin (2013a).
Cooper et al. (2002a) ofrecieron niveles crecientes de urea hasta 1,2% de la dieta y logra- ron 1,85 kg/día de aumento de peso en con 1,2% de urea versus 1,7 kg/día en el tratamiento con- trol (sin urea). Utilizaron novillos de biotipo grande (Cuadro 39) y dietas de 82% de grano hú- medo de maíz. El nivel de 13% de proteína bruta, incluyendo 0,8% de urea, permitió cubrir los requerimientos proteicos para superar 1,8 kg/día.
Pordomingo et al. (2003) compararon dietas a base de grano entero de maíz (73%) y ha- rina de girasol (15%) (T1, T2 y T3) con una dieta (T4) que incluyera harina de pescado (2,5%), y harina de plumas (3%) para aportar proteínas de baja degradabilidad ruminal (Cuadro 40). La adición de las fuentes proteicas de baja degradabilidad no mejoró el aumento de peso (Cuadro 41). Se detectó una leve depresión del consumo que resultó en una eficiencia de conversión su- perior (Cuadro 42). Pero, se destacaron los aumentos de peso obtenidos (superiores a 1,5 kg/día) en animales jóvenes (200 kg de peso inicial y 340 a 359 de peso final).
El potencial de las dietas de alta energía que no incluyen fuentes proteicas de baja degra- dabilidad ruminal, pero que cubren los requerimientos de proteína metabolizable, se corrobora también en los trabajos de Elizalde et al. (2003a,b) (Cuadros 8 y 26), Pordomingo et al. (2002 ab, 2003, 2004) (Cuadros 12, 20, 22, 41 y 44). De manera similar, Koenig y Beauchemin (2013a) observaron ausencia de respuesta en aumento de peso, consumo o conversion de alimento en dietas con inclusión de oferentes de proteina verdadera de distinta degradabilidad y nitrogeno no proteico, comparadas con una dieta basada en 85% de cebada, con 13.5% de proteina bruta y sin adición de fuentes adicionales de proteína.
En un intento por mejorar el valor biológico de las proteína degradable en las dietas de alto grano, Pordomingo et al. (2003 y 2004) incluyeron taninos condensados en dietas basadas en grano de maíz entero y harina de girasol. El tanino forma asociaciones con las proteínas que las hace menos solubles y degradables en medio neutro (rumen) pero disociables en medio ácido (abomaso). Ello permitiría incrementar la oferta a nivel intestinal de fracciones proteicas prove- nientes de la dieta y evitar la degradación parcial de la proteína en el rumen, con un consecuente incremento de la eficiencia de la metabolicidad de la proteína.
En dos ensayos se detectaron mejoras en el ritmo de engorde atribuibles a la adición de taninos, aunque se discute la magnitud de las diferencias y la implicancia de otros efectos debi- dos a la inclusión de los taninos sobre la intensidad y frecuencia de consumo. El Cuadro 40 des- cribe la composición de las dietas utilizadas en el primer ensayo. Los cuadros 41 y 42 muestran los resultados. De manera similar, el Cuadro 43 muestra la composición de las dietas del segun- do ensayo y los cuadros 44 y 45 muestran los resultados.
Al margen de la inclusión de taninos, se destaca en el segundo ensayo citado el aumento de peso logrado con dietas de 45% de grano, consideradas de recría o de aumento intermedio. Ese grupo de tratamientos aumentó a casi 1 kg/día, sin diferenciarse del testigo (sin taninos) del grupo de tratamientos de alto grano (70%). Debe puntualizarse, sin embargo, que la calidad del heno de alfalfa, clave en el planteo de alta fibra, permitió generar dietas con digestibilidad supe- rior al 65% (Cuadro 43).
La extracción con prensa y solventes del aceite del poroto de soja y del girasol generan como subproductos la harina de soja (frecuentemente denominada pellet de soja) y la harina de girasol, respectivamente. La harina de soja es el concentrado proteico de origen vegental (45 a 48 % de PB) que mayor concentración de PB ofrece debido a su bajo contenido de cáscara y aceite residual (2 %). Adicionalmente, el peril de aminoácidos de esa proteina es muy completo por lo que le confiere un alto valor biológico. La harina de girasol es un concentrado proteico de menor tenor que el de soja (29 a 35%), pero muy utilizado en bovinos en toda la región donde se produce girasol.
El costo de la harina de girasol, inferior al de harina de soja y su disponibilidad lo con- vierten en el recurso proteico más elegido en feedlots para carne. En contenido de aceite es simi- lar al de soja, pero el contenido de fibra es el que hace variar la proporción de proteina. Estos subproductos participan de las raciones en cantidades mínimas necesarias para alcanzar las metas de concentración proteica compatibles con los requerimientos de los animales. La participación de la harina de soja en las dietas de engorde se ubica generalmente en el rango del 10 al 20% y la de la harina de girasol en el rango del 15 al 30%.
Santini y Paván (2002) demostraron que con incrementos de harina de girasol del 20 al 43% en una dieta a base de silaje de maíz se puede incrementar el aumento de peso a 1,00 a 1,180 kg/día en dietas que fueron de 12 a 18% de proteína bruta (Cuadro 38). Esa evidencia muestra también la alta capacidad de reemplazo o sustitución de la harina de girasol en mezclas con silaje de maíz.
La soja tostada o (desactivada) constituye otro excelente recurso proteico (34 a 38% de proteína bruta) y también energético por su aporte de grasa (20%) y carbohidratos (32%). El tostado o calentamiento de la soja desactiva el factor antitripsínico (factor que bloquea la enzima tripsina y compromete la digestión de las proteínas en el intestino delgado) por ser una proteína termo-sensible, pero el costo del proceso hace que su uso comercial sea muy poco frecuente en el feedlot argentino. Sin embargo, la soja cruda puede ser utilizada también en bovinos. El rumen tiene una buena capacidad de metabolización y desactivación del citado factor por lo que el bo- vino tolera mucho más que otras especies el consumo de soja cruda.
Felton y Kerley (2004) reportaron inclusiones de hasta 24% de soja cruda en dietas de 70% de grano entero de maíz (Cuadro 46) o basadas en grano y silaje de maíz (Cuadro 47). No se reportaron efectos negativos debidos a la adición de soja cruda, con respuestas en aumento de peso (Cuadros 48 y 49), aunque se comentaron tendencias a efectos depresivos de conversión. El agregado de aceite incrementó la oferta energética de la dieta en todos los casos. Del trabajo citado emerge también que la adición de 24% de soja cruda no sería recomendable debido al riesgo de deprimir la conversión y el consumo emergentes de un exceso de aporte graso (aceite de la soja), de proteína y de factor antitripsínico en la dieta, excedentes que pueden comprometer la fermentabilidad de la fibra, incrementar el gasto energético en la eliminación del exceso de nitrógeno dietario como de poner en riesgo la digestibilidad de las proteina en intestino delgado. El exceso de aceite y los compromisos digestivos habrían incrementado también la incidencia de diarrea en el citado ensayo.
Albro et al. (1993) reportaron mayores aumentos de peso al incluir soja cruda versus soja extrusada y, Aldrich et al. (1997) y Eweedah et al. (1997) indicaron que la inclusión de soja cru- da al 16% de la dieta no provoca trastornos sobre la digestibilidad de la materia seca o de la fi- bra. La información sugiere también que el aporte (limitado) de aceite de soja incrementa la producción de propiónico y la lipogénesis hepática (el propiónico es insumo precursor de la pro- ducción de grasa intramuscular) (Smith y Crouse, 1984). También se ha reportado que el aporte oleoso de la soja en proporción limitada (inferior al 5% de la dieta) puede incrementar la tasa de marmolado y de engrasamiento sin alterar la composición de la grasa (Brandt y Anderson, 1990; Huerta-Leidenz et al., 1991). Pordomingo et al. (2007c) encontraron las mejores respuestas en aumentos de peso y eficiencia de conversión en terneras a corral con dietas que incluyeron 10% de soja cruda molida y 8% de harina de girasol (Cuadro 58). El agregado de 18% de soja cruda no resultó perjudicial de la eficiencia de conversión, pero se detectaron heces más blandas y ma- yor incidencia de diarreas.
Sobre esos antecedentes, en un estudio posterior, Felice et al. (2013) exploraron la res- puesta animal a la inclusión de proporciones crecientes de soja cruda o tostada (desactivada), reemplazando a la harina (pellet) de girasol en dietas completas basadas en grano de maíz (Cua- dros 59 y 60). Las dietas fueron isoproteicas e isoenergéticas, con una oferta creciente de aceite (en función del incremento de la proporción de poroto de soja incluido). Se diseñaron para cubrir los requerimientos proteicos de novillitos en crecimiento (NRC, 1996). La inclusión de soja tos- tada en reemplazo de harina de girasol en cantidades crecientes tendió a mejorar el ritmo de en- gorde sin afectar la conversión del alimento (Cuadro 60). La inclusión de soja cruda no afectó el aumento de peso y la conversión hasta el nivel de 18%. En el nivel de 24% de soja cruda se observó una depresión del ritmo de engorde promedio y una desmejora de la conversión (Cuadro 60). Dado que el incremento de la proporción de aceite en la dieta ocurrió en ambos casos, tosta- da y cruda, se atribuyó el efecto detrimental observado en el nivel 24% de soja al factor anti- tripsínico. Este ensayo y los precedentes permiten concluir que la inclusión de soja cruda en las dietas no debería superar niveles del 18% en los planteos de engorde.
En la última década, la producción de burlanda (residuos del destilado de los granos) de maíz y de sorgo (wet distiller’s grains) para la producción de etanol ha generado un cambio im- portante en el tipo de concentrado proteico incorporado a las dietas en el EEUU y Canada. La utilización masiva de este recurso ha promovido intensa investigación sobre las formas de inclu- sión, y las implicancias sobre la calidad del producto y el ambiente. La reciente incorporación de plantas de destilado de granos en Argentina apunta a generar un suproducto semejante, el que necesariamente encuentra su uso en la alimentación animal en confinamiento.
Los destilados o burlandas pueden ser húmedos o secos y su composición final cambia con los procesos que los general. Tienen la característica de ser oferentes protéicos y también energéticos, con moderado aporte de fibra digestible. Su aporte proteico variable y depende del proceso de extracción. La burlanda de maíz ofrece proteina en un rango del 29 al 35% y la de sorgo del 35 al 40% (May et al., 2012; Uwituze et al., 2010). May et al. (2012) ofrecieron burla- da de maíz o de sorgo en proporciones de 15 y 30% de la dieta (base seca) sobre dietas de alta energía basadas en maíz procesado en copos (flakes) (Cuadro 69). No se detectó un efecto de dilusión de la oferta energética con la inclusión de las burlandas. La burlanda de maíz generó eficiencias moderadamente superiores. Se destacan los aumentos de peso obtenidos (Cuadro 70).
Uwituze et al. (2010) ofrecieron burlanda seca de maiz a vaquillonas sobre dietas de alto contenido de grano y heno de alfalfa o silaje de maíz como fuente de fibra (Cuadro 71). Los autores no detectaron una interacción con la fuente de fibra o efecto de la misma, pero detectaron una moderada reducción del ritmo de engorde al incluir burlanda y remover otras fuentes de pro- teína (Cuadro 72). Por lo que se ha concluido que estos recursos tendrían una densidad energéti- ca equivalente a la del grano de sorgo y la cebada, apenas por debajo del maíz. Por ello son tam- bién considerados también oferentes energéticos, con capacidad de sustitución de los granos (Meyer et al., 2013; Segers et al., 2013; Schoonmaker et al., 2010; Depenbusch et al., 2009 a,b), por lo que se los incluye frecuentemente en proporciones superiores a las necesarias para cubrir requerimientos proteicos solamente (dependiendeo de las relaciones de precio con los granos). Segers et al. (2013) compararon la inclusión de 25% (base seca) burlanda seca de maíz, con glu- ten feed y haria de soja en dietas basadas en 75% de silaje de maíz en planteos de recría a corral (Cuadros 73 y 74). La respuesta en aumento de peso entre las dietas fue similar entre las que incluyeron burlanda y harina de soja, levemente superior a la que incluyó gluten feed.
En este sentido, se han realizado numerosas experiencias en EEUU donde se incluyeron las burlandas hasta el 40% de la dieta sin afectar la conversión el aumento de peso, (Buttrey et al., 2012; Schoonmaker et al., 2010; Loe et al., 2006). Las proporciones de inclusión comunes se ubican en el rango del 15 al 40%. Schoonmaker et al. (2010) incluyeron burlanda húmeda de maíz hasta el 40% de la dieta (base seca) sobre dietas de alta fibra y de baja fibra, sustituyendo al grano de maíz y el concetrado proteico en el primer caso y al silaje de maíz y el concentrado en el segundo, sin afectar el aumento de peso y la conversión del alimento a peso vivo (Cuadros 61, 62 y 63). Buttrey et al. (2012) probaron la inclusión de 20% (base seca) de burlanda húmeda de maíz a dietas basadas en maíz rolado seco o procesado en copos (flakes) (Cuadro 64). Las res- puestas en aumento de peso fueron similares en ambos tipos de dieta. La inclusión de la burlan- do no redujo el aumento de peso o empeoró la conversión (Cuadro 65). La respuesta fue mejor en las dietas que se basaron en maíz procesado en copos (flakes).
Buttrey et al. (2012) ofrecieron entre 15 y 16% de burlandas secas y húmedas de sorgo o maíz en dietas sin heno o no muy baja fibra (Cuadro 66). No obervaron diferencias importantes debido a la fuete de burlanda, pero dectecaron un efecto positivo al agregar la de sorgo a la dieta con heno (Cuadro 67). Las eficiecnias resultaron similares al tratamiento testigo, sin burlanda. Buttrey et al. (2013) detectaron mejoras en la performance (aumento de peso y conversión) de animales alimentados inclusión de 35% de burlanda húmeda de maíz sobre dietas de grano maiz aplastado o en copos (Cuadro 68 y 69). Sin embargo, el rendimiento de res y el nivel de engra- samiento resultó levemente inferior.
Meyer et al. (2013), Segers et al. (2013) y Schoonmaker et al. (2010) sugieren que la uti- lización de las burlandas en cantidades altas (superiores al 40%) puede generar alguna depresión del aumento de peso, respecto de dietas sin burlanda. Este efecto sería atribuible a la leve dilu- sión energética de la dieta que tal inclusión podría generar. Es conveniente conocer el contenido de azufre de la burlanda, ya que por el proceso químico de destilado y limpieza, es un subproduc- to enriquecido en azufre. El aporte de azufre podría ser excesivo y eventualmente tóxico si no se realizan ajustes en la oferta mineral y la cantidad de burlanda ofrecida es alta (superior al 30%).
También se han generado burlanda seca de triticale (36,8% de proteina bruta , 32.5 % FDN y 6 % de almidón) (Wierenga et al., 2010) y de trigo (38 % de proteina bruta, 33 % FDN y 1.8 % de almidón, 3.7% extracto etéreo, 0.11% Ca, 0.93% P, y 1.02% S) (Yang et al., 2012). Los autores incluyeron hasta 30% de burlanda seca de triticale o de trigo, respectivamente, sin depre- sión de conversión y aumento de peso (Cuadro 75). La adición de burlanda permitió reducir has- ta eliminar la cantidad de silaje incluido en la dieta como fuente de fibra (Cuadro 76).
El gluten feed, es otro subproducto resultante de la industria del maíz que ha incrementa- do su relevancia en la alimentación a corral deo bovinos en el área de influencia de algunos polos industriales. La presentación del gluten feed puede ser húmeda o seca, dependiendo de la planta que procesa el maíz y la estrategia de secado. Es un producto que puede considerarse proteico y energético (Segers et al., 2013; Loe et al., 2006) . El contenido de proteina se ubica en el rango del 16 al 28%. Valores frecuentes se ubican en el 23% (Loe et al., 2006). El contenido energéti- co se ubica en el rango del 75 al 80% del de maíz. Por el agregado de azufre al proceso, debería verificarse el contenido de azufre del producto y el resultante total de la dieta para no generar riesgos de intoxicación. Para que estas ocurran, la inclusión de gluten feed en la dieta debería superar el 50% (en base seca).
La inclusión en niveles de 15 a 35% es recomendada cuando se lo adiciona como oferen- te proteico, pero frecuentemente se intenta incluirlo en cantidades mayores para utilizarlo como oferente energético (Loe et al., 2006; Segers et al, 2013; Macken et al., 2004). Experiencias en EEUU (Kampman and Loerch, 1989) (Cuadros 79, 80, 81 y 82) han ofrecido gluten feed hasta alcanzar el 60% de la dieta, sin generar depresion del aumento de peso. Cantidades del 80% generaron aumentos de peso menores y peor conversión. Macken et al. (2004) demostraron que la inclusión de gluten feed húmedo de maíz hasta el 35% de la dieta no afectaría el aumento de peso y la eficiencia, comparado con proporciones menores o con un control a base de maiz en copos (extrusado y expandido), urea y concentrado proteico.
Loe et al. (2006) detectaron incremento de la respuesta animal en aumento de peso con la inclusión de gluten hasta el 52% de la dieta (base seca), basada en grano de cebada. En el mismo ensayo se evaluó el nivel de 35% de inclusión en una dieta basada en maíz aplastado seco. Esta superó en aumento de peso y conversión a las basadas en cebada (Cuadros 77 y 78). Farran et al. (2012) indicaron que la inclusión de 35% de gluten feed humedo (en base seca) a dietas de terminación basadas en maíz aplastado, aporta proteina, energía y un nivel de fibra que permite de la reducción de la fracción de heno de pastura o su eliminación sin comprometer la fermenta- ción ruminal, incrementar riesgo de acidosis, o afectar el aumento de peso y la conversión. Par- sons et al. (2007) evaluaron la inclusión de gluten feed de maíz al 40% (base seca) con la reduc- ción hasta la eliminación del heno de alfalfa (Cuadro 83). La inclusión de gluten habría permiti- do la reducción de la fuente de fibra sin resultar en depresión significativa del consumo o la con- versión (Cuadro 84).
Vander Pol et al. (2009) ofrecieron gluten feed de maíz al 20% (base seca) a dietas basa- das en grano húmedo de maíz o con inclusión de burlanda seca con solubles (al 40% base seca) y no detectaron diferencias en la respuesta animal (aumento de peso y conversión). El gluten feed, combinado con la burlanda de maíz permitó reducir la cantidad de grano molido de maíz del 67 al 27%, sin perder potencial de engorde. La experiencia se realizó con animales pesados de frame grande y los aumentos de eso superaron los 2 kg/día.
Nitrógeno no proteico
La inclusión de urea (o formas similares de nitrógeno no proteico) en dietas de rumiantes requiere de alta capacidad fermentativa en rumen y un actividad ruminal desarrollada, de lo con- trario se convierte en un tóxico que puede generar amoniosis (intoxicación por circulación de amoníaco en sangre). Fluharty et al. (2000) demostraron que terneros de 3 meses al destete tole- ran dietas de alta energía con un contenido de urea de 0.4% de la dieta (base seca).
Dietas con alto contenido de urea son comunes en la experimentación (Pordomingo et al., 1999a,b, 2000). En novillos, Cooper et al. (2002a) demostraron que con la adición de urea hasta el 1,2% de la dieta la respuesta en aumento de peso fue lineal (Cuadro 39). La evidencia experimental indica que la respuesta a la inclusión de urea existe en la medida quel se logra y mantiene un balance de proteína metabolizable positivo y siempre que no aparezcan limitaciones en la oferta de energía al rumen. Por su parte el aporte de nitrógeno dietario es efectivo hasta que se cubre la necesidad ruminal (Koenig y Beauchemin, 2013b).
En novillos o vaquillonas por encima de los 250 kg de peso vivo, los niveles máximos de urea ofrecidos sin problemas de intoxicación o costos energéticos adicionales (detoxicficación) se ubican en el 1,2% de la dieta total. Niveles de 0,8 a 1,0% de urea son comunes en muchos feedlots. A manera de seguridad, es conveniente llegar al nivel máximo progresivamente, empe- zando con un 0,5% de base e incrementando la cantidad de urea a razón de 0.1% cada 2 o 3 días. Ese acostumbramiento es importante para adaptar por un lado al comportamiento ingestivo y el metabolismo del animal (el rumen, el hígado y la recirculación de la urea), y por otro al mezcla- do y manejo de la dieta.
El agregado de urea exige también un buen mezclado con el alimento, y sería mejor si se incluye en un pelleteado con grano y harinas proteicas o afrechillo que participen de la dieta. En el caso de no contar con la posibilidad de incorporarla en un concentrado proteico peleteado se puede agregar perlada o granulada en una premezcla con grano molido, que la diluye en un ma- yor volumen de material, el que luego se incorpora al total del alimento.
En dietas húmedas a base de silajes de granos húmedos o de planta entera, el mezclado y distribución homogénea de urea y sales minerales es más fácil de lograr. La humedad favorece la adherencia de las partículas finas y secas. La urea es muy higroscópica por lo que tiende a absorber agua y adherirse, disolviéndose en parte. El mojado de granos secos con 5 a 8% de agua previo a la inclusión de la urea y el núcleo mineral o sales es una práctica frecuente en sis- temas de mucho grano entero y poca fibra. En estos casos, si se incorpora harina de girasol, soja, o afrechillo de trigo peleteados es conveniente mojar el grano previo a la carga de estos concen- trados. Los pellets de estos subproductos son higroscópicos y se deshacen si entran en contacto con el agua, perdiendo consistencia y palatabilidad.
La máxima homogeneidad de mezclado de ingredientes que se aportar en cantidades in- feriores al 1% (urea, sales minerales y vitaminas) se logra mediante inclusión en forma líquida, asperjando sobre los ingredientes mayores (granos, harinas, silaje o heno). Ello exige sin embar- go, de una mezcla y dilusión previa a la carga del mixer o camión de reparto. Las mezcladoras estáticas de alto volumen son poco utilizadas en los feedlots de Argentina pero serían la mejora herramienta para el mezclado adecuado previo al reparto. Por su costo se justifican en empren- dimientos grandes (más de 5000 animales).
Requerimientos de proteina
El requerimiento de proteína bruta y metabolizable decrece con el incremento de la edad, del peso y del nivel de engorde del animal (Figura 1). Terneros al destete (más de 5 meses) tie- nen requerimientos de 14 o 16% de proteína bruta y los novillos de más de 400 kg del 11 a. 13%. El requerimiento de proteína bruta, sin embargo, depende también de la metabolicidad de la pro- teína. Si la calidad de la proteína es baja y una fracción alta de la misma (superior al 35%) pro- viene de una fuente nitrogenada no proteica, los requerimientos de proteína bruta se incrementan para alcanzar los mínimos de metabolizable. En esos casos podemos encontrarnos que un terne- ro requiere 16% de proteína bruta y un novillo en terminación necesitar un 14%.
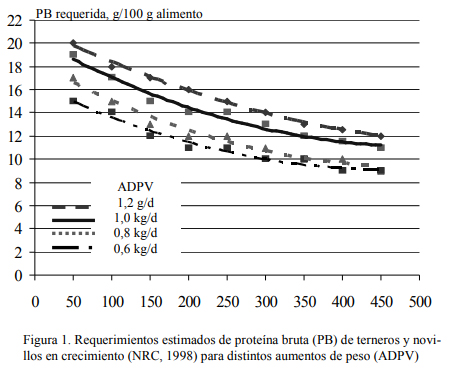
La historia nutricional previa al ingreso a los corrales de engorde también tiene efectos sobre los requerimientos proteicos y la respuesta a la proteína de la dieta (Sainz et al., 1995). El requerimiento de proteína se incrementa con el aumento compensatorio en el animal. Recrías muy restrictivas generan efectos compensatorios en la fase de corral, donde los requerimientos proteicos se elevan en 1 o 2 puntos (ej. 14 a 16 % en terneros y de 12 a 14 % -base seca- en novi- llos).
Desde el punto de vista de la calidad también es mayor el requerimiento en animales jó- venes donde el aporte de formas nitrogenadas no proteicas (ej. urea) debería ser tal que el su aporte equivalente en proteína bruta no supere el 33% del total de proteína bruta ofrecida. En terneros de destete precoz y anticipado (60 a 120 días) no sería conveniente incorporar fuentes de nitrógeno no proteico (ej. urea) debido a la limitada capacidad de producción de biomasa micro- biana en el rumen.
Minerales y vitaminas
Para garantizar la conversión de alimento a carne en estos planteos no debe obviarse o subestimarse el rol del suplemento mineral y vitamínico para evitar carencias y deterioro de la conversión. Debe asegurase en ese suplemento la presencia de sal común, calcio, fósforo y mag- nesio, sumamente necesarios para animales jóvenes en crecimiento. Dada las dificultades de mezclar cantidades pequeñas, se sugiere utilizar correctores minerales que ya contienen sal y macro-minerales (Ca, P, Mg) pre-mezclados con los micro-minerales y la monensina u otro ionó- foro. Estas observaciones son muy importantes en los casos en que se plantee no utilizar ningún tipo de fibra larga (henos).
Los animales grandes, próximos a la terminación (ej.- más de 380 kg en razas británicas) tienen en términos relativos menos requerimientos de minerales y vitaminas que los terneros o vaquillonas y podría evitarse el uso de minerales y vitaminas si la fase de terminación no supera los 60 días. En categorías más jóvenes (terneros y vaquillonas) el uso de estos elementos es inevitable ya que los requerimientos superan los aportes de minerales que pueden hacer los gra- nos y los concentrados proteicos por sí solos.
En el caso del fósforo, el aporte de fósforo a través del grano puede ser suficiente (Erick- son et al., 1999, 2002). Los requerimientos de fósforo de acuerdo a NRC (1996) asumen un requerimiento diario de mantenimiento de 16 mg/kg peso vivo y de 3,9 g por cada 100 g de pro- teína retenida, con una absorción media del 68%. Sin embargo, Erickson et al. (1999 y 2000) han probado que los modelos actuales sobredimensionan los requerimientos reales, pudiendo reducirse entre un 40 a un 50% sin afectar factores de crecimiento o terminación. Ello implica que de formulaciones con 0,35% de fósforo (de acuerdo a tablas NRC, 1996), podría formularse con 0,16 a 0,22% de fósforo). El Cuadro 50 muestra los resultados de uno de los ensayos citados (Erickson et al. (2000) donde se pone en evidencia la insensibilidad de la respuesta al incremento de fósforo en la dieta de animales alimentados a corral. El aumento de peso resultó semejante entre tratamientos, también la eficiencia de conversión y las características de la carne (espesor graso, tamaño de ojo de bife y marmolado).
En estas categorías, el nivel de calcio y magnesio requiere ser incrementado por encon- trarse siempre en cantidades deficitarias en las dietas de alto grano (Kincaid, 1988). Los reque- rimientos de calcio tabulados por NRC (1996) exigen una concentración de al menos 0,35% de la dieta (base seca).
Erickson et al. (1999) detectaron una menor eficiencia en dietas con 0,70% de calcio, comparadas con las de 0,35%. De manera similar, Dowe et al. (1957) observaron un deterioro del aumento de peso cuando el Ca se incrementó de 0,4 a 2,6%. Varner and Woods (1972) ob- servaron una mejora lineal en el aumento de peso cuando contenido de Ca incrementó de 0,20 a 0,41% , pero no detectaron mejoras cuando se superó el nivel del 0,50% en novillos sobre dietas de alto concentrado energético. Huntington (1983) alimentaron con 0,3, 0,6, 0,9, y 1,2% de Ca en la forma de carbonato de calcio y observaron que la concentración óptima de Ca se encontraba entre 0,3 y 0,6% en dietas de novillos que recibían una dieta con 85% concentrado. En dietas con aporte graso (que contienen aceites o grasa animal), la interacción entre la grasa y el calcio en exceso podría resultar en una depresión de la absorción de grasas y una menor ganancia de peso (Bock et al., 1991.)
Ricketts et al. (1970) evaluaron 3 relaciones entre calcio y fósforo (Ca/P): 1:1, 4:1 y 8:1 con terneros y concluyeron que la performance era inferior en los que fueron alimentados con la relación 8:1. Wise et al. (1963) evaluaron rangos de 4:1 a 14,3:1 con terneros de 114 kg calves and y concluyeron que los óptimos de performance se dieron entre los rangos 1:1 a 7:1. Erickson et al. (1999) sugieren como la relación de 4:1 como relación de referencia para las formulaciones con alto contenido de grano.
El uso de conchilla (carbonato de calcio) es común como fuente de calcio pero de escaso valor biológico. Existen en el mercado concentrados minerales comerciales con fuentes de cal- cio y fósforo de mayor valor biológico. También se utilizan oxidos de calcio y sulfatos de calcio. El fosfato dicálcico es una fuente común de fósforo y calcio. Las formas orgánicas de presenta- ción de los minerales (quelados) son de mayor disponibilidad biológica y por lo tanto mas efi- cientes, pero su costo y disponibilidad las hace frecuentemente inviables en el pais.
El aporte de zinc, cobre, selenio, molibdeno, cobalto, hierro, boro, y manganeso es nece- sario (Hill et al., 1996). En particular, los dos primeros son frecuentemente deficitarios en mu- chos suelos y aguas de la mayor región ganadera del país. Estos elementos están íntimamente relacionados con la integridad de los epitelios y membranas, la funcionalidad del hígado y los procesos de deposición de proteína y hueso en el animal. Se requiere de su suplementación en todos los planteos intensivos.
El Cuadro 50 muestra el detalle de los ingredientes minerales incluidos y la oferta mine- ral resultante en dos dietas similares. Comúnmente los núcleos vitamínicos se diseñan para cu- brir los requerimientos minerales de las raciones de confinamiento suponiendo un aporte mínimo de los ingredientes mayores (granos, harinas), pero en función de estos puede diseñarse el aporte mineral de la dieta con mayor precisión. En las tablas de nutrición (NRC, 1998, 2000, 2002) se muestran los requerimientos para las distintas categorías y tipo de animal. En algunos casos y en las categorías de mayor peso el aporte de minerales requerido por encima de los aportados por los granos, harinas y heno o silajes puede ser mínimo o nulo. La sal estará siempre en déficit.
El agua tiene incidencia en el aporte global de micro-minerales. Carencias de cobre, zinc y hierro en el agua pueden requerir de un aporte adicional de estos elementos. Por su capacidad para secuestrar calcio, cobre, magnesio y zinc, las aguas con alto contenido de flúor (mayor a 0,7 ppm) arsénico (mayor a 0,3 ppm) y sulfatos no serían recomendables. La dilución de esas con- centraciones será la primera opción. Ante la ausencia de alternativas habría que incrementar la oferta de calcio, magnesio y micro-minerales en la dieta. Pero, dicha corrección será siempre parcial y poco efectiva.
Ionóforos y protectores de la fermentación
La acidosis es el principal problema del engorde a corral con alto contenido de grano. Por su rápida fermentación, el almidón produce acidificación del rumen (Owens et al., 1996). Se incrementa el contenido de ácido láctico (la producción de ácido aumenta y su transformación a otras formas disminuye) en rumen y cae el pH. También aumenta la absorción de ácidos hacia la circulación sanguínea. El animal entra en un estado acidótico y comatoso. Se acalambra la mus- culatura ruminal y la circulación periférica se hace más lenta. Se pierde elasticidad en el sistema circulatorio y muscular, particularmente el venoso se ve muy afectado y se edematizan los teji- dos.
Se genera edema e inflamaciones de epitelios y membranas de los tejidos periféricos (muy visibles en las extremidades). El animal camina con dificultad y con dolores, y se producen heridas en la piel y en las articulaciones. Las lesiones en las patas pueden derivar en enfermeda- des podales com el pietín. La menor motilidad ruminal promueve la retención de gases que se acumulan en el rumen e hinchan al animal del lado izquierdo, de manera similar al empaste, pero esta hinchazón (timpanismo) no es espumosa por lo que cede ante una punción del rumen que permita liberar el gas.
En el aparato digestivo la funcionalidad ruminal se ve afectada. Ocurre paraqueratosis e inflamación del epitelio ruminal, se erosiona la superficie de absorción, degeneran las papilas y pierden la capacidad de absorción, y se hieren las paredes del rumen. Las heridas evolucionan en llagas, vías de ingreso de bacterias, proceso que deriva en infecciones hepáticas y abscesos. Los animales con acidosis subclínica retardan el crecimiento y el engorde. Esos animales tienen me- nor eficiencia ruminal, y menor función hepática, causales de un consumo deprimido y una me- nor conversión.
La presencia de ionóforos (monensina, lasalocid, etc.) en la dieta es necesaria para evitar el sobre-consumo y la acidosis o empacho (Pordomingo et al., 1999 a,b; Raun et al., 1976). El más utilizado es la monensina. Monensina es un ionóforo monovalente que opera como protec- tor del sobre-consumo y modula la fermetabilidad de las proteínas y la producción de metano. La regulación del consumo y el mantenimiento de la función de las paredes del rumen son fun- damentales para evitar caer en un síndrome acidótico.
El modo de acción de los ionóforos es múltiple. En primer lugar afectan las poblaciones bacterianas en el rumen. Promueven una fermentación de mayor captura de energía en formas de ácidos débiles más reducidos (propiónico vs acético (Pordomingo et al., 1990; Santini y Di Mar- co, 1983; Church, 1988). Reducen la metanogénesis (formación de metano – gas), la tasa de proteólisis ruminal y la población de protozoos. Intervienen en el balance de algunos minerales y en su absorción en las bacterias del rumen (afecta la bomba de sodio y potasio) (Pordomingo et al., 1990). Reducen además las poblaciones de coccidios y la degradación de vitaminas y pro- vitaminas (Duff et al., 1990).
A través de todos estos efectos combinados, se aumenta la eficiencia de uso de la dieta, se homogeniza y regula el consumo, y se reduce el riesgo de acidosis subclínica. Se han deter- minado mejoras en la conversión entre el 8 y el 12% y eliminación del 100% del riesgo de acido- sis con la adición recomendada y rutinaria de monensina en las dietas de alto contenido de grano en feedlot (Suber y Bowman, 1998; Pordomingo et al., 1999b). Niveles de 0,75 a 0,100 g de monensina por cada 100 kg de peso vivo serían los recomendados para controlar los factores de riesgo citados en dietas con consumos de 2,8 a 3% del peso vivo (base seca) en las dietas con escasa o nula fibra larga (menor al 10%).
Expresado en proporción del alimento a razón de 29 a 31 mg de monensina por kilo de alimento total (29 a 31 gramos en 1000 kg). En el caso en que esté formulada al 10% (ej. Ru- mensin), se agregará a la dieta incorporando entre 0,75 y 1 gramo de rumensin por cada 100 kg de peso del animal o 290 a 310 gramos por cada 1000 kg). En el caso de uso de lasalocid (Bova- tec) la proporción de ingrediente activo es de un 15% mayor que la de monensina para lograr efectos similares en aumento de peso (Pordomingo et al., 1999a). Se debe recalcar que estas sugerencias suponen una dieta de base de alta calidad (con energía para alto ritmo de engorde) y cantidades bajas de fibra larga.
En dietas basadas en silajes de maíz, sorgo, avena o pasturas (más de 30% de silaje en base seca) el riesgo parálisis ruminal y de acidosis es menor por lo que la dosis de monensina pueden reducirse a la mitad. Su uso, sin embargo, sería de utilidad por sus efectos sobre la meta- bolicidad de la dieta, la modulación del consumo diario y en definitiva en la mejora de la conver- sión.
El acostumbramiento
El acostumbramiento merece especial atención. En ese período el rumen del animal de- berá acostumbrarse progresivamente a fermentar altas cantidades de almidón sin que se provo- quen trastornos digestivos. El rumiante proveniente del pastoreo no está preparado para fermentar y digerir grandes cantidades de almidón (Church, 1988). El rumen tiene que adaptarse, tanto la microflora ruminal (bacterias del rumen) para realizar el trabajo fermentativo, como la funcio- nalidad de las paredes del rumen y el hígado del animal para remover y procesar los metabolitos (nutrientes) emergentes de la fermentación.
Por un lado, la microflora ruminal debe mudar de preferentemente celulolítica (especiali- zada en degradar celulosa) a amilolítica (especializada en degradar almidón). Este cambio de dominancia de poblaciones bacterianas es disparado en la presencia de almidón y para alcanzar un nivel de actividad amilolítica alto y sostenido lleva entre 10 y 14 días (Storry y Sutton, 1969). El almidón expuesto en el rumen se fermenta rápidamente degradándose a ácidos grasos de dis- tinta longitud de cadena y actividad ácida, los deseables son los ácidos más débiles (pK alto comparado con el del ácido láctico, 4,7 vs 3.8). Para que el proceso fermentativo sea estable y sostenido, estos metabolitos deben ser removidos del medio, previa absorción a través de las paredes del rumen (Church, 1988).
Si por motivos fermentativos (ritmo de fermentación muy alto) o funcionales (absorción comprometida) la remoción de ácidos es deficiente, la acidez del rumen aumenta (el pH cae), lo que reduce a su vez la absorción y promueve una mayor depresión del pH. En ese escenario los ácidos grasos volátiles no removidos continúan su degradación hacia formas ácidas más fuertes (ej. ácido láctico). La presencia de ácidos en alta concentración lastima o erosiona la superficie de absorción (las paredes del rumen). El mismo medio ácido impide el desarrollo de las bacte- rias metano-genéticas (generadoras de metano, una de las formas más reducidas del carbono, que opera como mecanismo de apoyo en la reducción de potencial reductor) (Cheng et al., 1998).
Paralelamente, se reduce la motilidad ruminal por efecto del aumento de la concentración de ácido láctico en la musculatura de las paredes ruminales. La pérdida de motilidad resulta en la reducción drástica de la eruptación y la acumulación progresiva de gases en el rumen. El animal progresa hacia un cuadro de timpanismo por empacho. La sobrecarga de ácido láctico en circu- lación supera la capacidad detoxificadora del hígado y aumentan los niveles en la circulación periférica comprometiéndose los tejidos epiteliales y articulaciones en primer lugar (efectos de infosura o hinchazón y deformación en patas y manos, dolores reumáticos y complicaciones motrices) (Cheng et al.,1998) .Cambian también la química sanguínea y los perfiles endócrinos (Brown et al., 2000a).
Por otro lado, la capacidad de absorción del epitelio ruminal tiene que adecuarse a un mayor ritmo de remoción de ácidos grasos volátiles. Con la fermentación amilolítica crece la producción total de ácidos grasos y crece la proporción de propiónico en relación a la de acético. También aumenta la carga de ácido láctico en el licor ruminal y en la circulación sanguínea del tejido de las paredes del rumen. El epitelio ruminal (las papilas) tiene que adaptarse a ese medio más ácido e incrementar su actividad metabólica. Esta adaptación requiere de tiempo y un cam- bio de pH moderado. Si la caída del pH es rápida y decrece en pocos días por debajo de 5.0 el proceso adaptativo se detiene y comienza una reacción auto-protectora del epitelio pero degene- rativa del mecanismo de absorción (paraqueratosis).
El proceso de acostumbramiento de los animales a la dieta de alto contenido de almidón necesita de 14 a 21 días. En ese período el rumen se adapta a la nueva fuente de energía. Esto implica que las paredes ruminales adecuan su estructura papilar para soportar un medio más áci- do, que la maquinaria de remoción de ácidos grasos se estimula, y que el hígado aumenta la ca- pacidad de procesamiento de metabolitos ácidos en formas tolerables para el resto de los tejidos (ácidos grasos de cadena larga y triglicéridos).
Por su parte, el comportamiento animal cambia, en ese período se de reducir progresiva- mente el nivel de estrés por encierre permanente y aprender rutinas. Durante el acostumbramiento el animal aprende a comer con mayor frecuencia y menor intensidad. El estrés retarda el acos- tumbramiento a dietas de alta energía, particularmente por los cambios en la circulación sanguí- nea que provoca. Altera el comportamiento ingestivo, provoca asincronías en el tracto digestivo y altera los procesos digestivos.
Importantes en la regulación de la acidez (o el pH) son la salivación y le reciclado de electrolitos via la pared ruminal. Mediante la saliva se produce la incorporación de sustancias “buffer” y es la principal via (Waldo, 1976). La producción de saliva está directamente relacio- nada con la rumia o re-masticación del alimento y esta necesidad íntimamente asociada a las fracciones gruesas de la dieta, particularmente la fibra larga (Church, 1988). Debería comenzar- se con una dieta mayoritaria en heno (fibra) para en el término de 14 a 21 días estar en la dieta de alto grano. La velocidad de este proceso depende de la rutina y la capacidad de adaptación de los animales.
Existen varias estrategias de acostumbramiento o adaptación sugeridas en la experimen- tación o practicadas por las empresas. Lyle et al. (1981) reportaron que el reemplazo de heno por grano a razón de 0,5 kg/día les permitía alcanzar el consumo de una dieta de 90% concentrado en 14 días. Pordomingo et al. (1999 a,b) reportaron una estrategia de acostumbramiento a dietas de 90% concentrado consistente en 3 etapas, una primera donde el heno constituía el 60% de la dieta durante 7 días, una segunda con 50% de heno durante 7 días y una tercera con 25% de heno durante 7 días, para a partir del día 21 se pasaba a una dieta con 90% concentrado y 10% de fi- bra.
Pordomingo et al. (1999ab, 2002, 2003) sugieren un acostumbramiento en dos etapas pa- ra llegar a dietas de 75% de grano. Se propone ofrecer un alimento con 50% de grano en una primera etapa durante 7 días, luego incrementar a 65% de grano en los 14 días subsiguientes (segunda etapa), para incrementar al nivel de 75% de grano correspondiente a la dieta final. En dietas con más de 75% de grano sería necesario anexar una tercera etapa de una semana de ali- mento basado en 75% de grano para, cumplida la misma pasar al nivel mayor y definitivo. Una estrategia simple consiste comenzar con una dieta con 30% de grano durante 4 días, luego pasar a 40% de grano durante 4 días, luego a 50% de grano durante 4 días , luego a 60% de grano du- rante 4 días y finalmente instalarse en la dieta de mayor concentración (ej. 75 a 85% de grano).
Como alternativa al suministro de dietas de distinta concentración energética en etapas, algunos feedlots han implementado con éxito un sistema de dietas de alta y baja fibra combina- das durante los efrecidos en el día. Se comienza con una dieta de alta fibra (60% al menos) du- rante 7 días, luego se ofrece por la mañana la mitad de la cantidad diaria con la misma dieta de alta fibra y por la tarde la de baja fibra. Esta estrategia se sigue por 10 a 14 días subsiguientes y a partir de la tercer semana (21 días) se ofrece la dieta de baja fibra (terminación). Esta estrate- gia simplifica la construcción de dietas diversas en el mismo día y se simplifica la logística de adimistración de dietas a los corrales.
Para lograr buena adaptación a las rutinas, los comederos y el entorno se prefiere dar prioridad a la calidad de alimentos durante la etapa de adaptación. Ello implica que en las dietas de alta fibra se ofrescan henos o ensilajes, granos y concentrados proteicos de buena palaptabili- dad, dejando los granos de menor aceptación y henos o silajes de menor palatabilidad para las etapas posteriores.
En el caso de animales con síntomas de acidosis u otro trastorno digestivo es conveniente reducir la concentración energética de la dieta mediante el incremento del contenido de fibra (ej. pasar a una dieta de 40 o 50% de heno y 50% de concentrado), pero no pasarlos a una dieta de heno solo porque el animal no tendrá un rumen acostumbrado a digerir fibra. El heno debería ser de alta calidad. Luego de recuperado el nivel de consumo y desaparecidos los síntomas acidóticos (inflamaciones, hinchazón, edemas, dificultades motrices, etc.) se le podrá incrementar el nivel de concentrado a los niveles anteriores, aunque a estos animales sería conveniente no expo- nerlos a dietas de muy alto grano (ej. superiores a 75% de grano). Muchos de ellos tienen una susceptibilidad innata a los trastornos digestivos debida, entre otras causas, a una menor capaci- dad de salivación o menor motilidad ruminal.
El acostumbramiento es más dificultoso en terneros que en novillitos y novillos, espe- cialmente si los terneros nunca han estado expuestos al grano, provienen de un destete anticipado o de muy bajo peso, tienen un estado general pobre (se encuentran delgados y deficientes en vitaminas y minerales). El método de acostumbramiento puede ser cualquiera de los arriba suge- ridos. Sin embargo, aquí es importante la calidad del heno utilizado durante al menos los prime- ros 14 días. Ese heno tiene que ser de buena calidad y sería conveniente que durante los dos primeros días los terneros coman una dieta con 75% heno de buena calidad (ej. heno de alfalfa).
El consumo y la forma de alimentación
El consumo es el primer factor y el más directamente asociado al crecimiento y al au- mento de peso. Altos consumos en forma sostenida (mayores al 2,5% del peso vivo) se correla- cionan con altos aumentos de peso (NRC, 2002). El nivel de consumo diario voluntario de bovi- nos para carne sobre dietas de alta calidad se aproxima al 3% del peso vivo (observar consumo en cuadros más adelante). En las categorías jóvenes el consumo será equivalente al 2,8 a 3,2% del peso vivo o algo superior. En las categorías más grandes (novillos de 350 kg para arriba) el consumo diario puede variar entre el 2,6 al 2,8% del peso vivo. En términos absolutos, un novi- llo de 300 kg de peso vivo estaría dispuesto a comer entre 8 y 9 kg de materia seca en alimento total por día. Los terneros, en relación a su peso, comen más que los animales de mayor edad, por lo que en un terneros de 200 kg de peso podría esperarse un consumo ad libitum de 3% de su peso o superior, o sea 6 a 6,5 kg de materia seca/día.
Con dietas de alta concentración de energía metabolizable (superiores a 80% de concen- trado), el consumo puede deprimirse con respecto a los niveles logrados con dietas de menor concentración energética debido al efecto de los mecanismos quimiostáticos sobre la saciedad. Por su parte, con el incremento del contenido de componentes fibrosos (henos, silajes y cáscaras) por encima del 50% de la dieta, también el consumo voluntario diario puede deprimirse. En el caso de dietas basadas en silaje de planta entera (con alto contenido de agua y pH por debajo de 4,5) debe tenerse en cuenta que se puede deprimir el consumo respecto de los forrajes secos, siendo frecuente consumos de menos de 2,5% del peso vivo.
Debemos tener en cuenta las consideraciones sobre el consumo para no subestimarlo o desconocerlo. Si se pierde la noción del consumo, perdemos el control del indicador más rele- vante del engorde a corral: la conversión (de alimento a aumento de peso). Si bien muchos feed- lots trabajan ofreciendo a voluntad (comedero lleno), sería ideal alimentar en forma intermitente varias veces al día (2 o más) para evitar grandes volúmenes en el comedero que pierden calidad, la separación de las fracciones componentes del alimento por selección o por migración de las partículas más finas hacia el fondo del comedero, y tener un mayor control diario.
La oferta que promueve un consumo frecuente per en cantidades bajas (baja intensidad) permite estabilizar el pH ruminal en niveles superiores a 5.5 y eliminar los efectos indeseables debidos a la caída del pH por consumo de gran cantidad de alimento en ofertas menos frecuentes (Kaufmann, 1976; Kaufmann et. al., 1980). Este es en parte el objetivo de muchos métodos de procesado del alimento que aportan volumen y humedad al alimento para reducir la tasa de con- sumo de energía metabolizable, maximizando el uso de la misma.
En la práctica, los feedlots de gran escala alimentan tres veces al día. En esos sistemas se sugiere comenzar con la primera oferta de alimento de 4:00 a 6:00 de la mañana, realizar la segunda entre las 11:00 y las 13:00 de la mañana, y la tercera entre las 16:00 y las 18:00. En los casos en que resulte muy poco práctica o costosa esa frecuencia de alimentación, se propone ofrecer el alimento dos veces por día al menos, dividiendo en 2 la oferta diaria (según los cálcu- los sugeridos anteriormente). Al ofrecer 2 o 3 veces al día el consumo será más homogéneo, la separación de componentes de la dieta menor, habrá menor incidencia de acidosis subclínica (empacho) y también menor desperdicio que alimentando una vez al día. Aún con alimento disponible en el comedero, la oferta de alimento fresco promueve el consumo intermitente (co- mer un mayor número de veces) y reduce la tasa de consumo y reduce el estrés.
Para mantener un rumen en buen funcionamiento, sin acidosis subclínica o cambios drás- ticos en la generación de metabolitos que pudieren alterar la función hepática, sería aconsejable que el procesado de los granos, el mezclado de la dieta y la forma de alimentación permitan con- tar con un alimento completo y balanceado frente al animal cada vez que este se acerque al co- medero (Russell y Rychlik, 2001).
La oferta de fracciones por separado (concentrado por una lado y rollo o fuentes de fibra por otro) no es conveniente. Sin embargo, en el caso de hacerlo por no contar con otra posibili- dad de aporte de fibra, se sugiere dar heno (rollo o fardo) por horas (2 o 3 horas por día), permi- tiendo el acceso a los rollos por ese período de tiempo y luego retirar los animales con un hilo eléctrico. De esta manera, se pueden colocar varios rollos por corral para que todos los animales puedan tener acceso a algo de rollo, poder ofrecer un rollo de calidad media como para que todos los animales estén dispuestos a comer y lograr un consumo más homogéneo en el lote, pero evi- tar un consumo excesivo mediante la oferta por tiempo limitado.
Se sugiere planear el comedero con un frente de 30 cm por animal, dispuesto sobre una de las caras del corral (acceso de los animales por un lado solamente). Esto permitirá que al menos el 70% de los animales tengan acceso al comedero al mismo tiempo. Los comederos de bolsa o plástico funcionan bien para planteos temporarios o de baja escala. (Ver instalaciones en capítulos siguientes.)
La rutina de alimentación es importante. No deberían alterarse o molestarse los animales con pesadas demasiado frecuentes u otros movimientos innecesarios. Toda pesada o vacunación debe ser planeada tratando de evitar ayunos prolongados que puedan exponer a sobre consumos y empachos posteriores.
Alimentación restringida
La alimentación restringida al 85 o 90% del consumo voluntario del animal es una estra- tegia que algunos feedlots incorporan para mejorar la eficiencia de conversión y regular los so- bre-consumos (Galyean, 1999). En la mayoría de las dietas de alta energía (a base de 70 a 90% de grano), la restricción de la oferta diaria de alimento en un 10 a 15% con respecto al consumo voluntario esperable resulta en aumentos de peso similares a los alcanzables en consumo ad libi- tum, particularmente en etapas tempranas del engorde (Hicks et al., 1996). Se favorece incluso el flujo de compuestos nitrogenados hacia el tracto inferior (Murphy et al., 1994 b; Sip et al., 1991).
En la medida en que se aproxima la fase final y el grado de engrasamiento deseable es elevado, los sistemas de consumo sin restricciones superan en tasa de aumento y engrasamiento a los que restringen el consumo (Rossi et al., 2000; Loerch y Fluharty, 1998b). El Cuadro 51 resume resultados del ensayo comparativo de Rossi et al. (2000), con una dieta ofrecida ad- libitum, de alto grano, con la misma ofrecida al 90% del consumo voluntario. Los aumentos de peso resultaros similares y la eficiencia de conversión mejoró con la restricción de la oferta al 90%.
A niveles de engrasamiento bajos (ej como los demandados en Argentina), el grado de engrasamiento y el rendimiento resulta similar para ambos sistemas (Galyean, 1999). Pero, de- bido a un consumo diario menor se logra una mejora en la eficiencia de conversión con la estra- tegia de alimentación con oferta restringida (Murphy y Loerch, 1994; Loerch y Fluharty, 1998a; Owens et al., 1997; Wessels y Titgemeyer, 1997). Este sistema obliga también a un mayor se- guimiento de los animales y a la “lectura permanente del comedero” para manejar la oferta sin excederse en la restricción. Rossi et al. (2000). La estrategia de alimentación para ganancias programadas es menos frecuente en Argentina que la de aumentos máximos, pero se impone en los sistemas que incorporan el engorde a corral de recría o de terminación a fecha fija.
Estrategias de recría o crecimiento a tasa limitada (ej. 0,7 a 0,8 kg/día) utilizan también a la restricción de la oferta diaria (a niveles de 65 a 70% del consumo voluntario o 2,0 a 2,5% de peso vivo), y la proporción de fibra en la dieta, como las dos herramientas para planteos de au- mentos de peso programados. Estos modelos son de fácil implementación cuando existe un re- curso fibroso como el silaje para manipular la concentración energética de la dieta y más comple- jo cuando la dieta es de alto grano. La restricción de la oferta diaria para controlar el engorde requiere de un mayor espacio de comedero para que los animales pueda comer en forma simultá- nea, de lo contrario el consumo puede ser desparejo.
En diseños de dieta de oferta restringida es conveniente dimensionar la oferta de proteína en términos absolutos ( gramos de proteina bruta consumida por día por el animal). La restric- ción al consumo en animales en crecimiento debería imponer restricciones en la oferta energética pero no limitar la satisfacción de los requerimientos proteicos para el crecimiento (aumento de peso) esperado. En otros términos, la oferta de energía podría limitarse a la mitad de la requerida para cubrir el potencial de crecimiento y engorde de animales jóvenes, pero no debería limitarse a la mitad la oferta proteica. Una restricción del consumo en un 30% para limitar el engrasa- miento temprano, dependiendo de los requerimientos del animal, implica una reducción de la demanda de proteina (sin comprometer el desarrollo normal) no mayor al 10%. En términos prácticos, los requerimientos de proteina bruta del novillo o vaquillona de las razas británicas y sus cruzas, dentro del biotipo mediano y los pesos de faena, común en la ganadería argentina, se mueven en el rango de los 800 a 1300 g/día. Los más jóvenes y livianos, con aumentos de peso controlados tienen demandas de proteina por debajo de 1 kg/día y los de mayor peso y en ritmo de engorde alto tienen por encima del misno. La calidad de la proteína permite un mayor ajuste de estos valores de referencia y un ahorro en cantidad diaria pero para ello se hace necesario tener información sobre su metabolicidad. El ajuste por metabolicidad permitiría lograr mejoras de la eficiencia de conversión de la proteína bruta y de la dieta entre el 10 y el 15% (Pordomingo et al., 2003; Cuadro 40, 41 y 42).
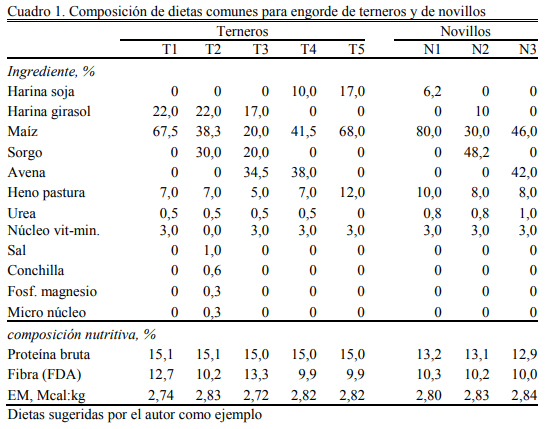
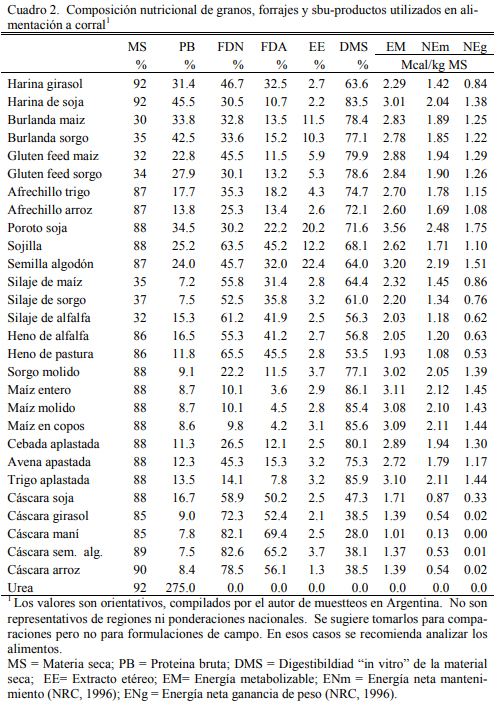
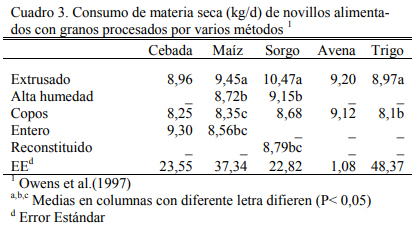
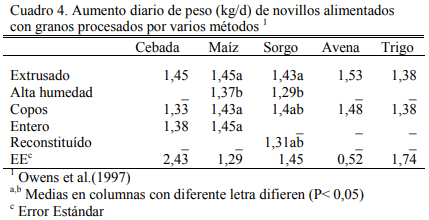
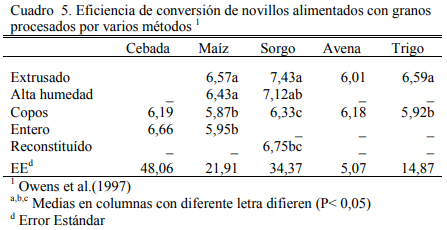
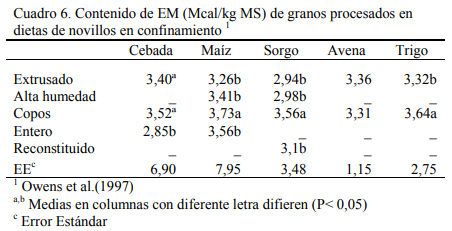
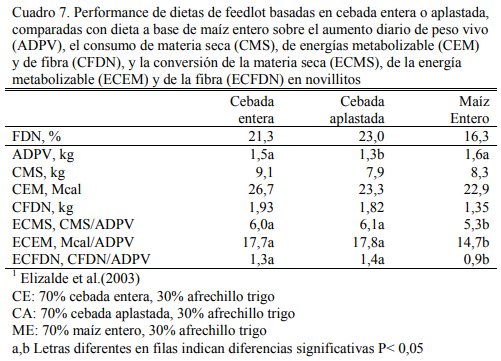
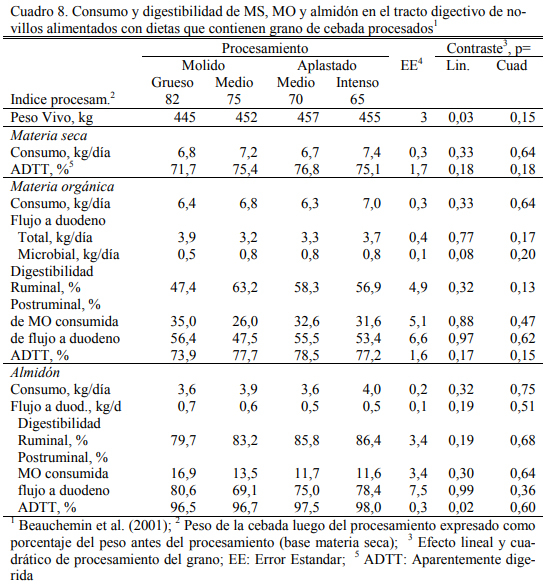
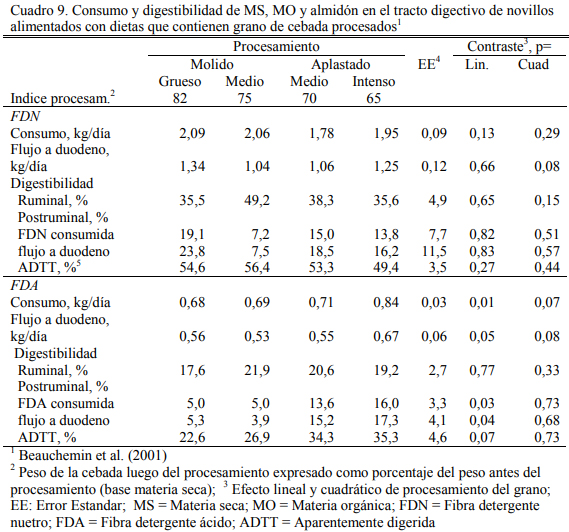
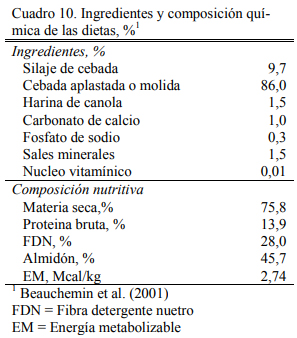
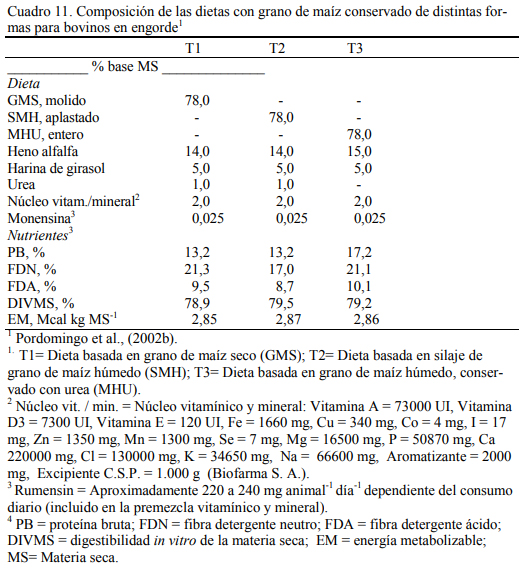
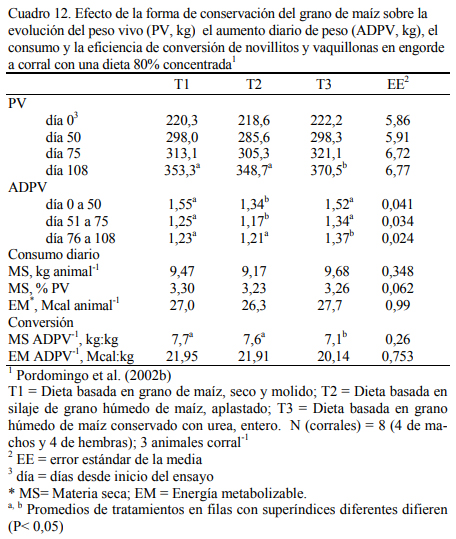
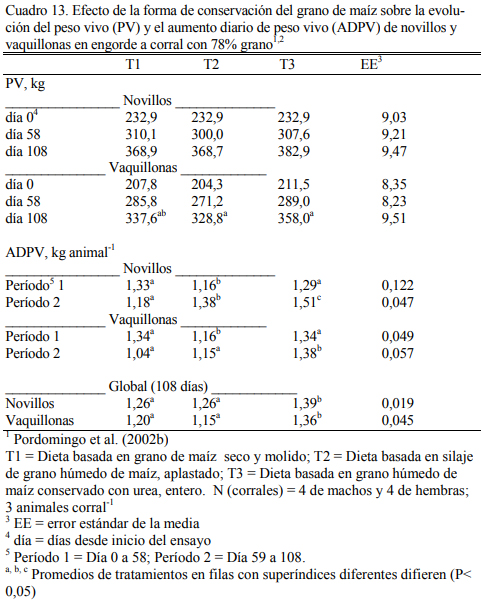
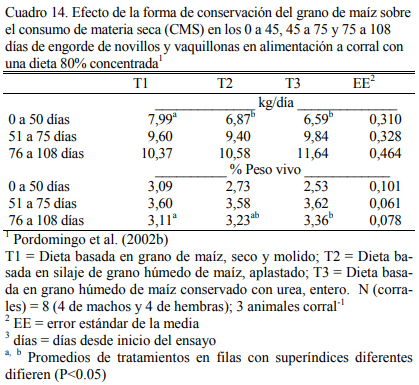
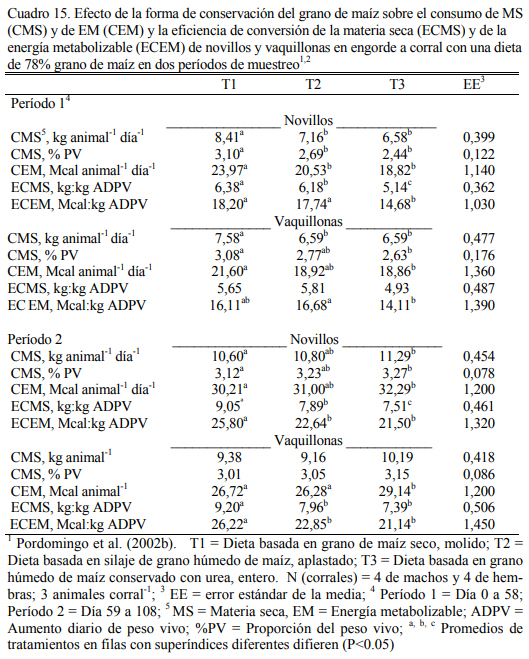
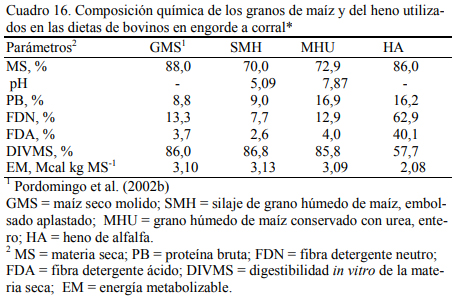
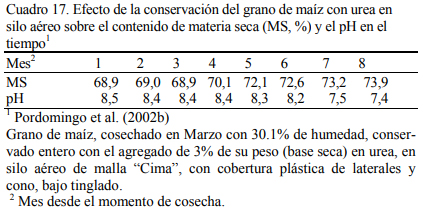
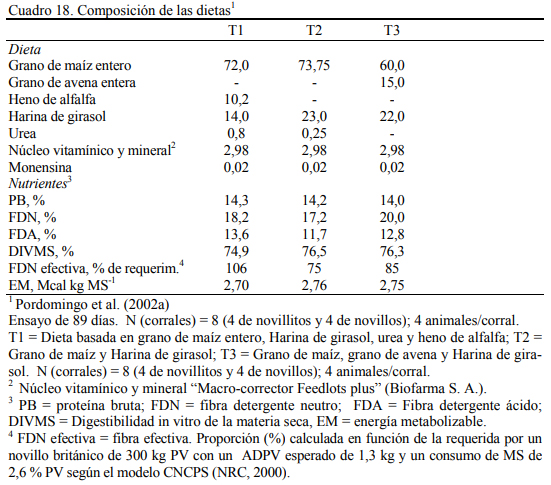
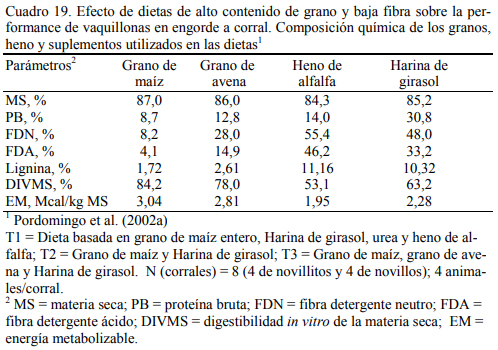
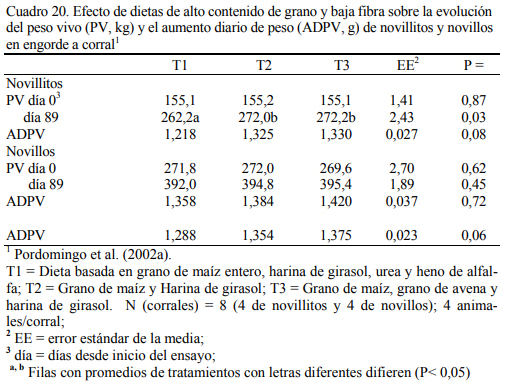
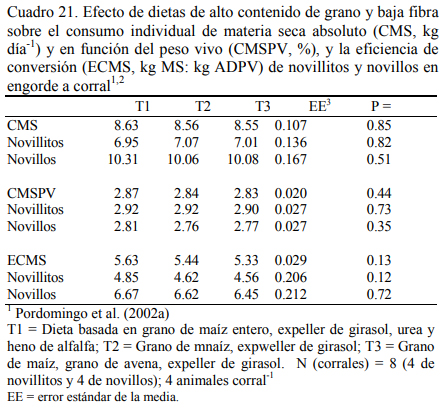
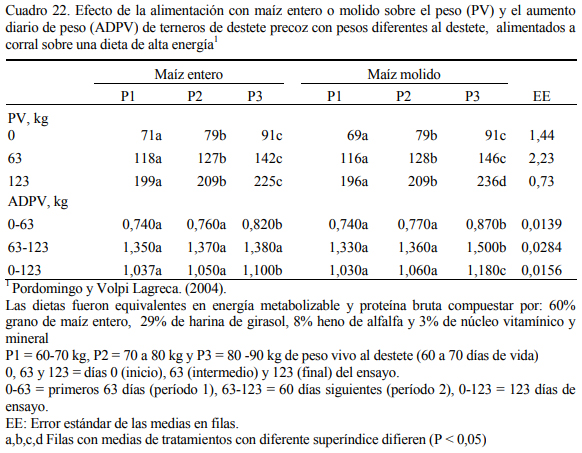
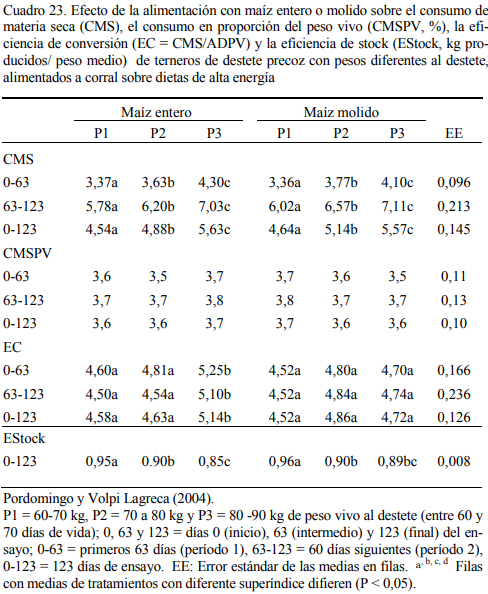
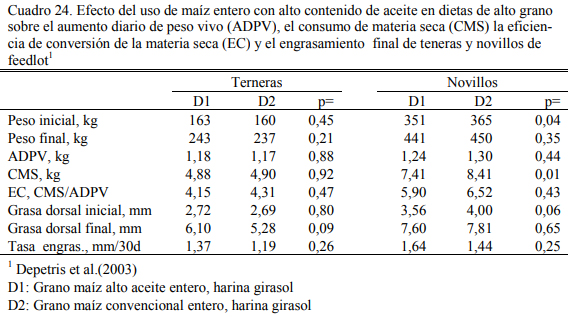
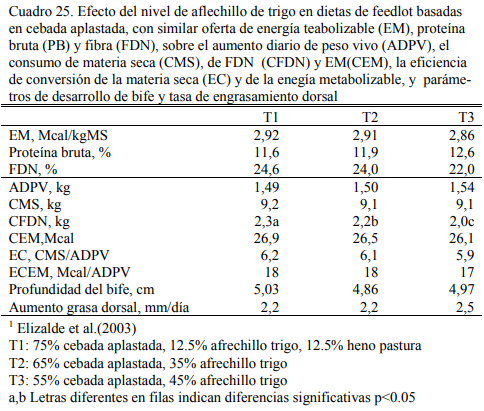
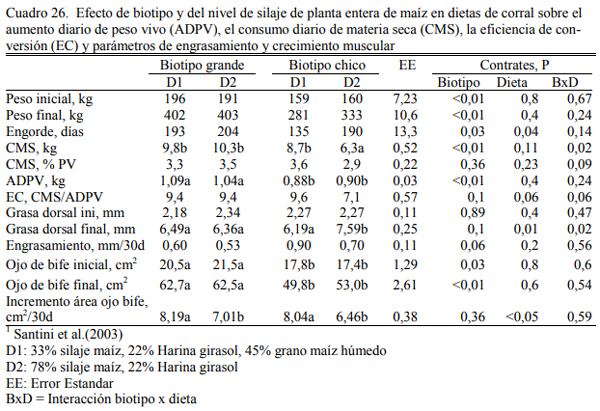
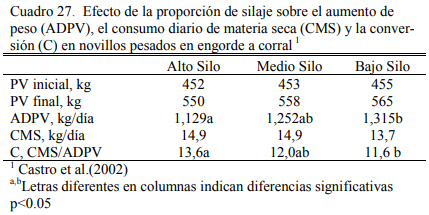
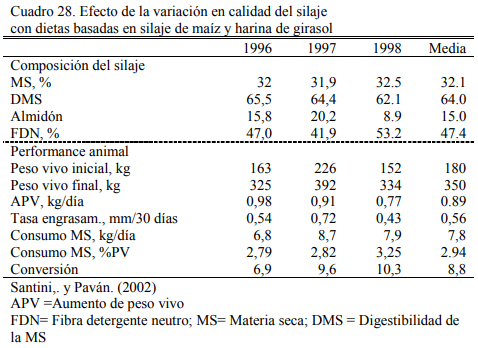
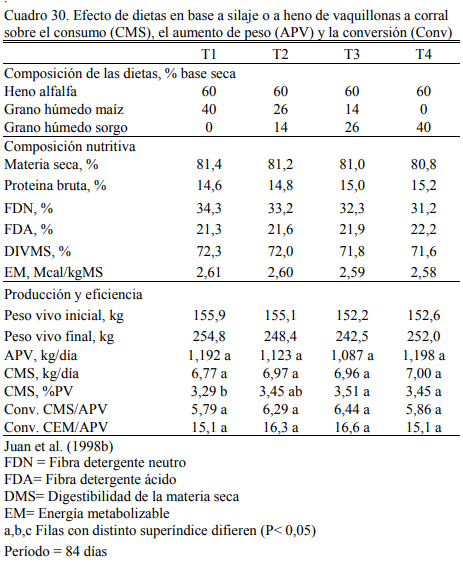
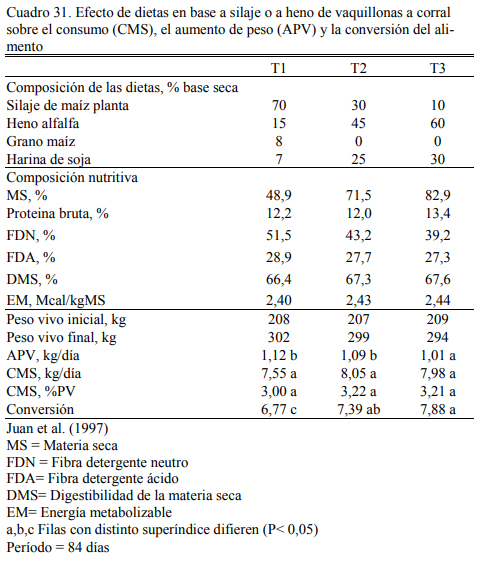
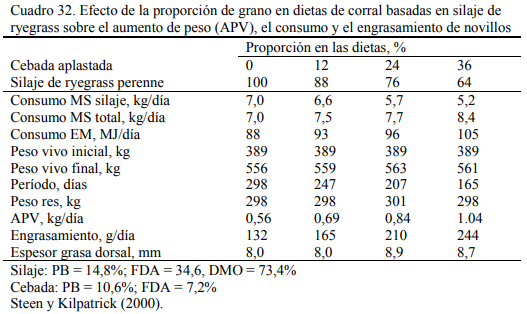
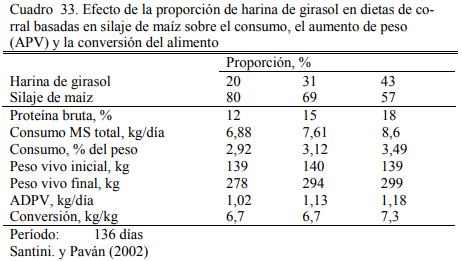
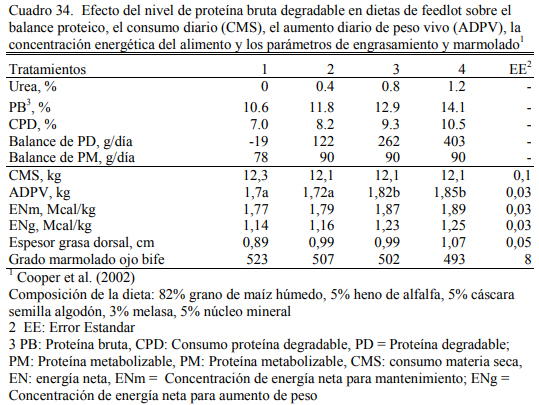
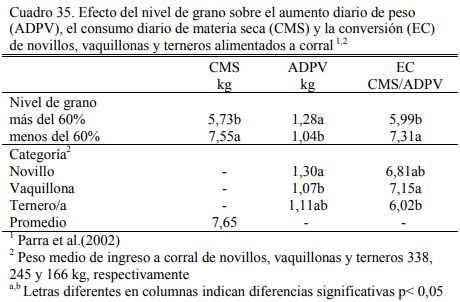
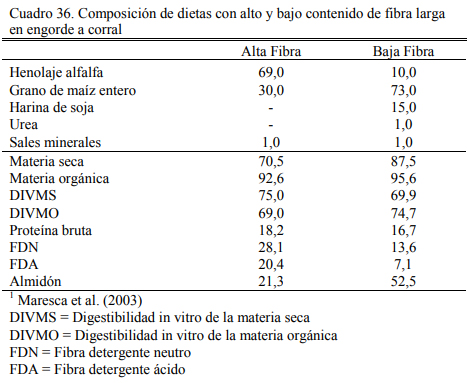
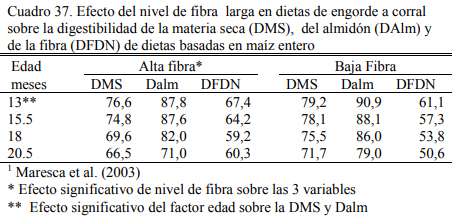
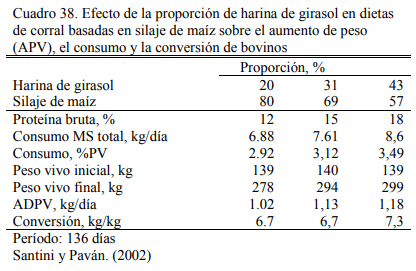
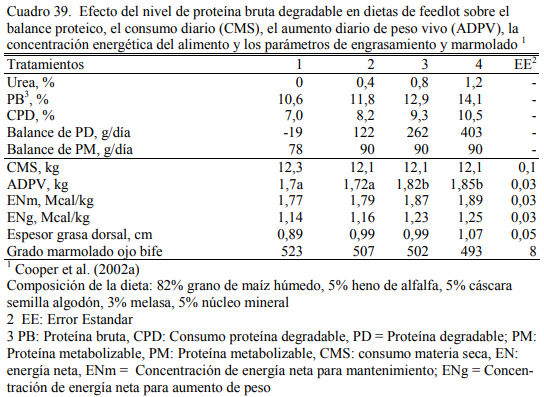
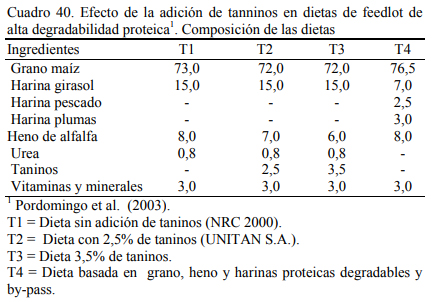
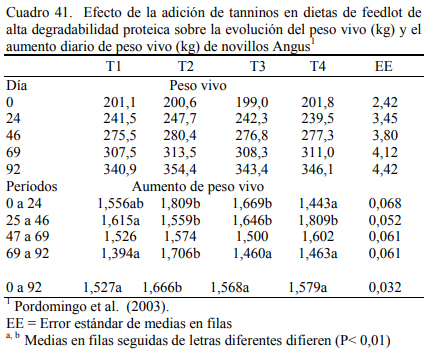
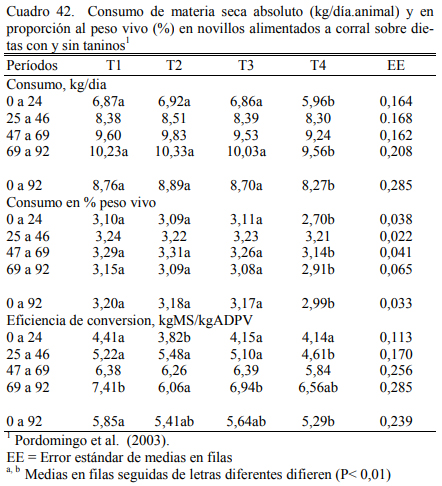
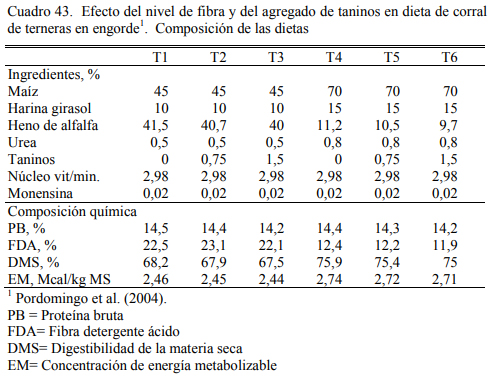
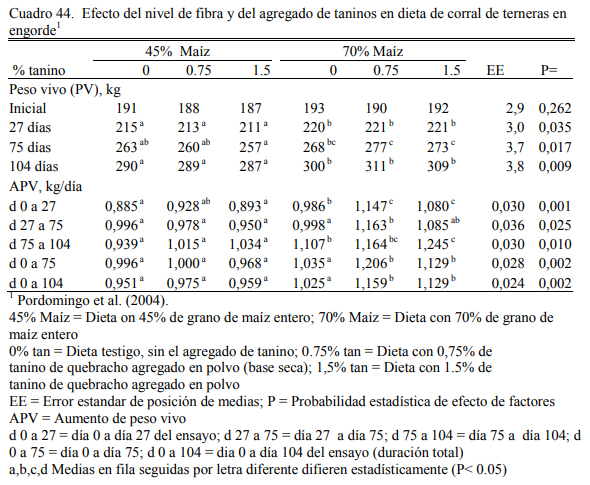
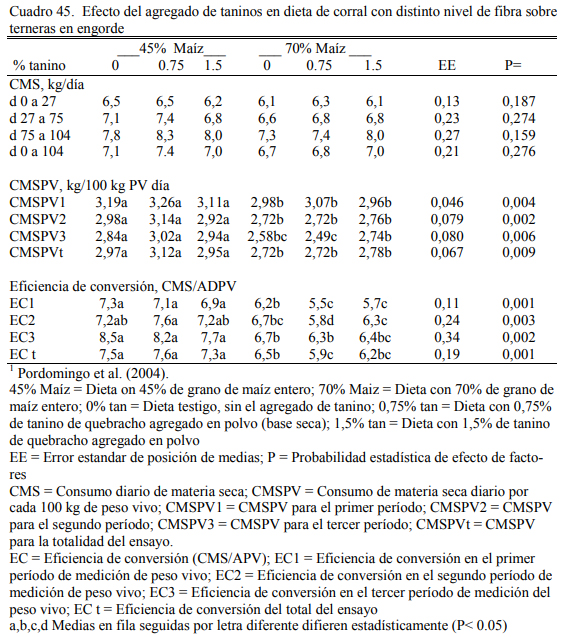
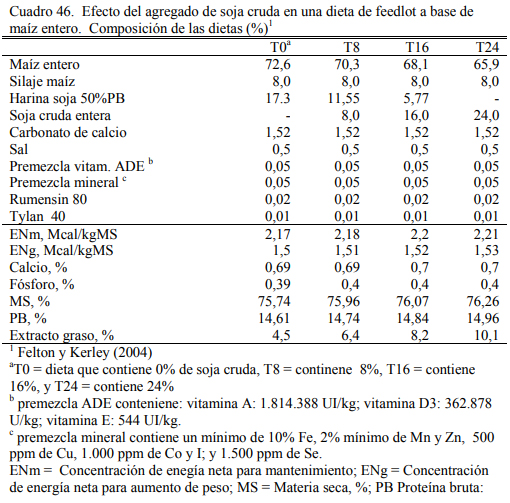
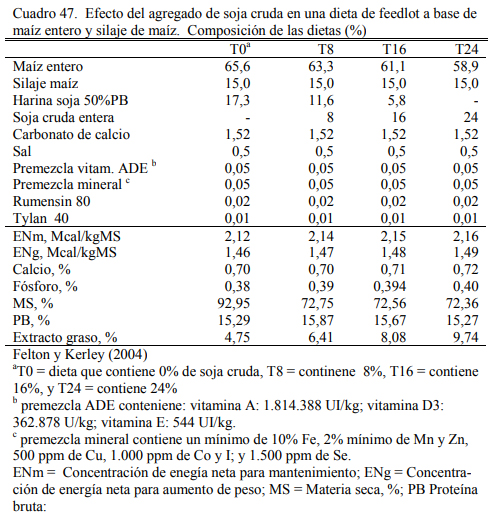
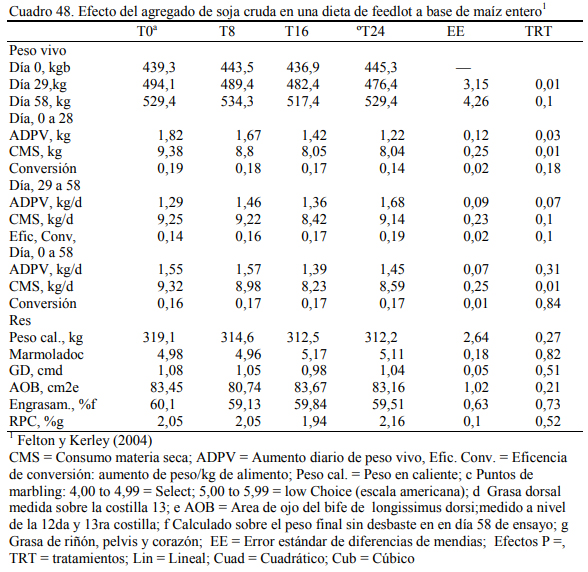
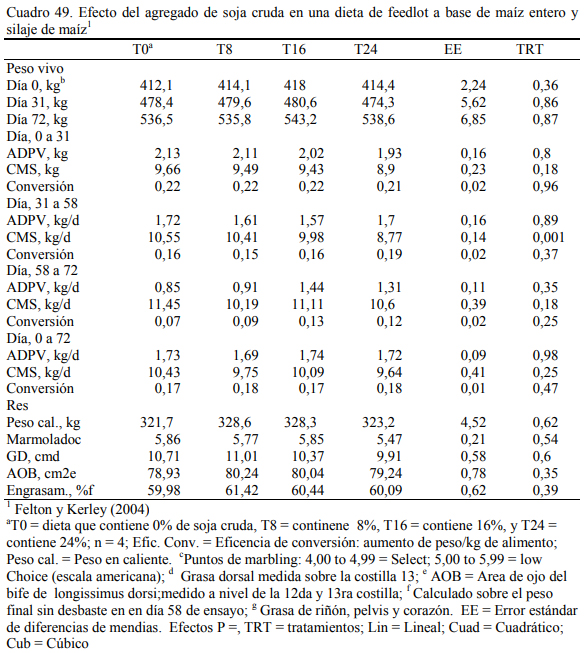
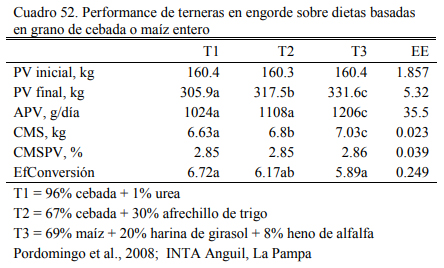
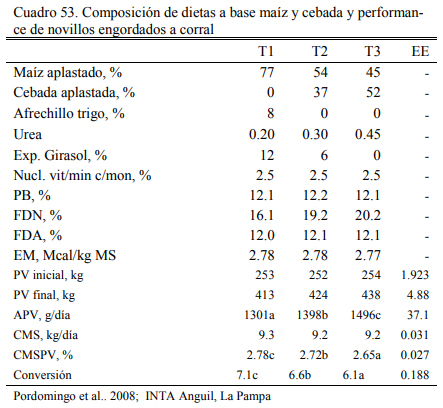
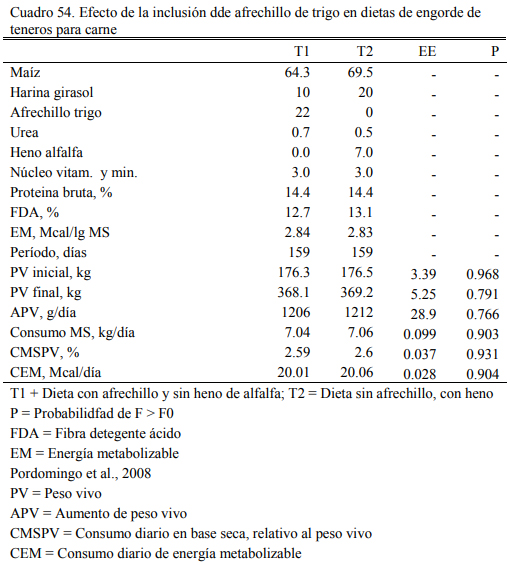
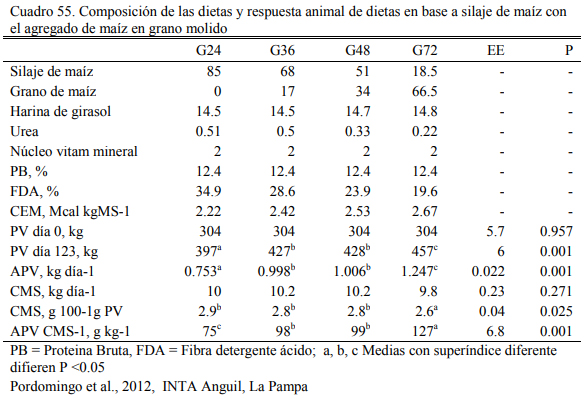
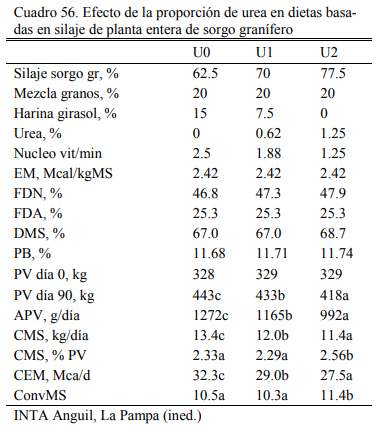
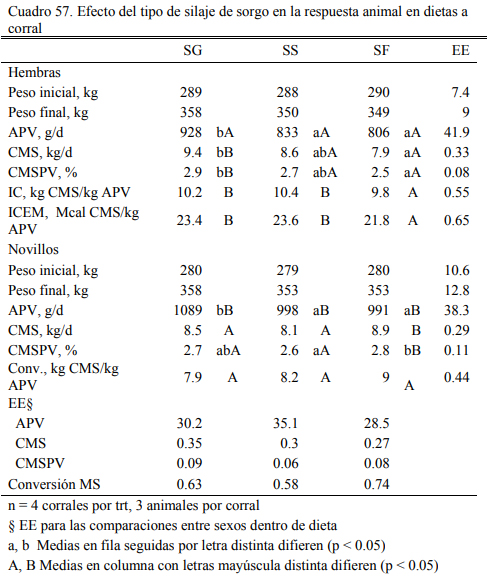
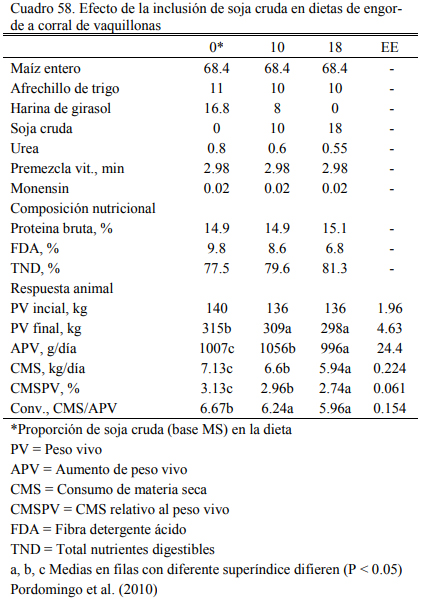
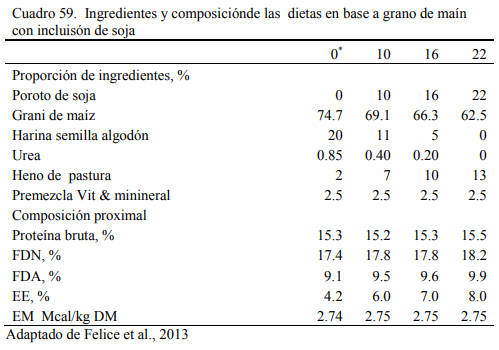
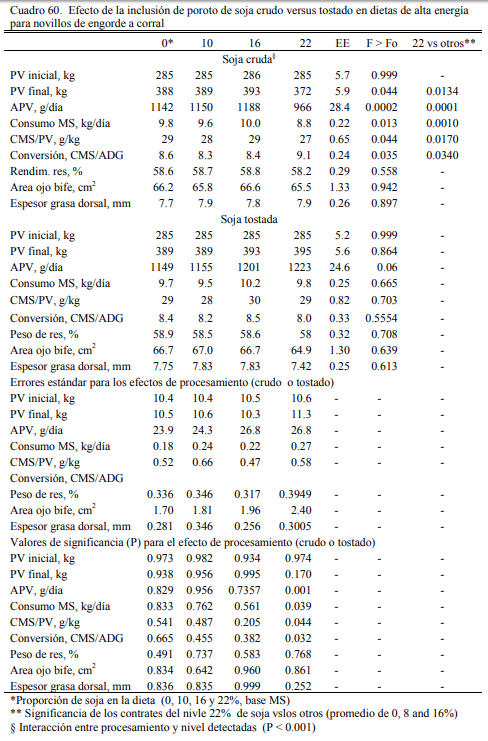
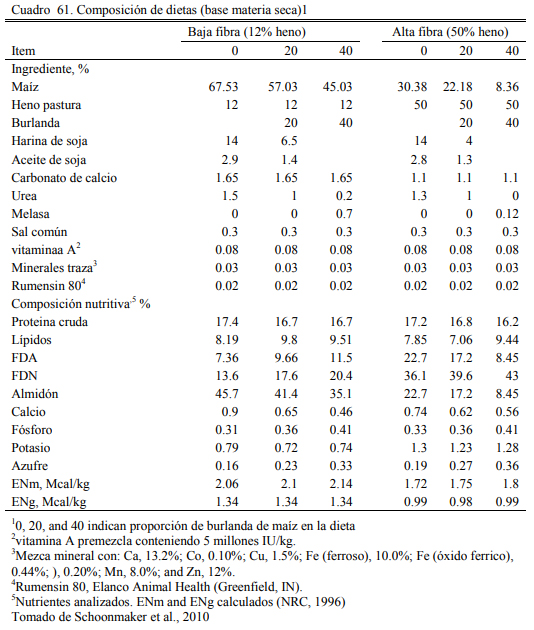
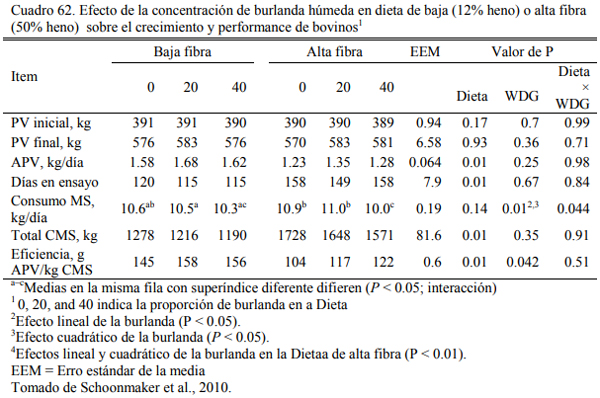
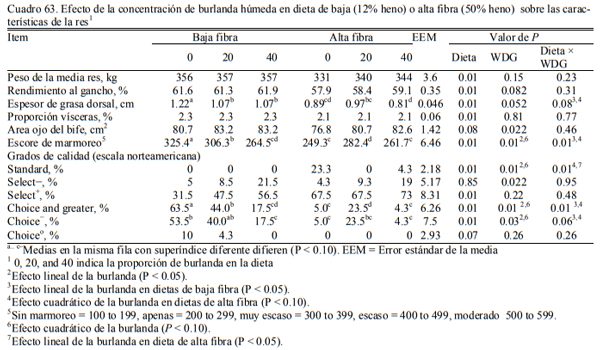
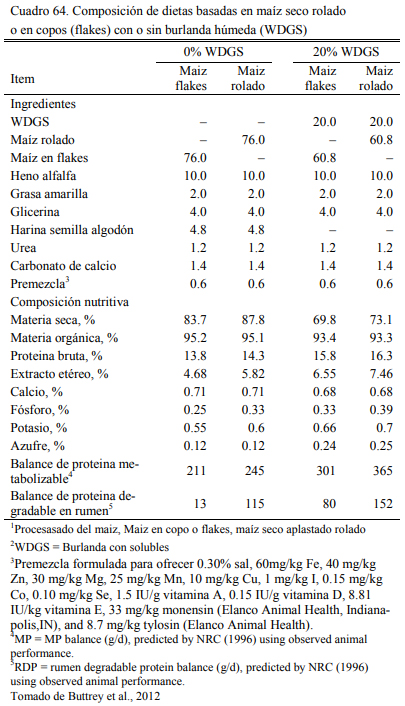
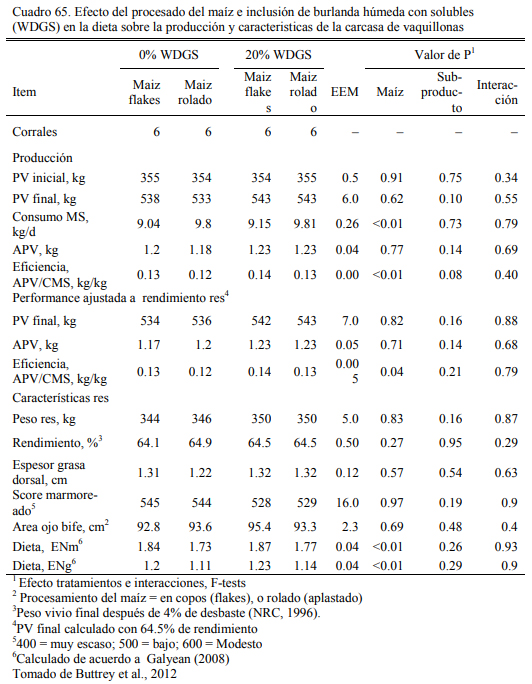
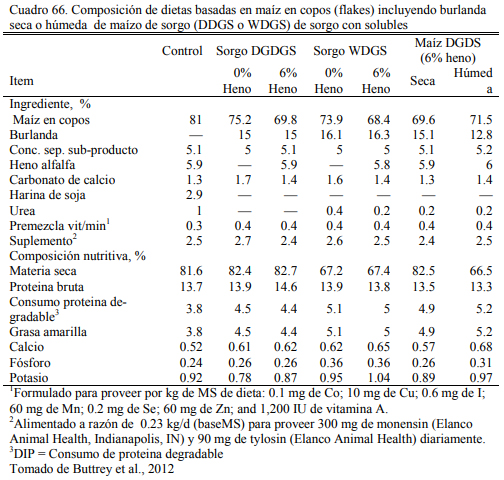
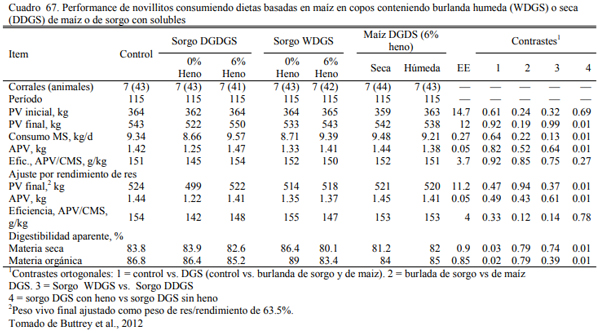
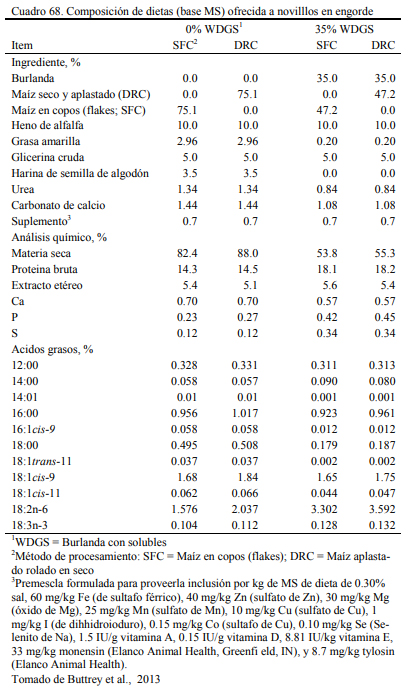
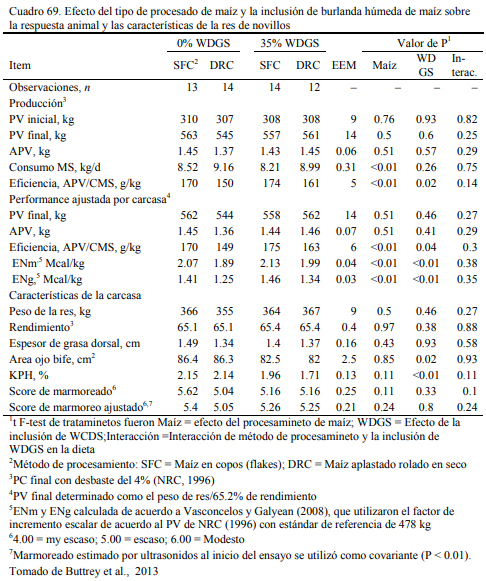
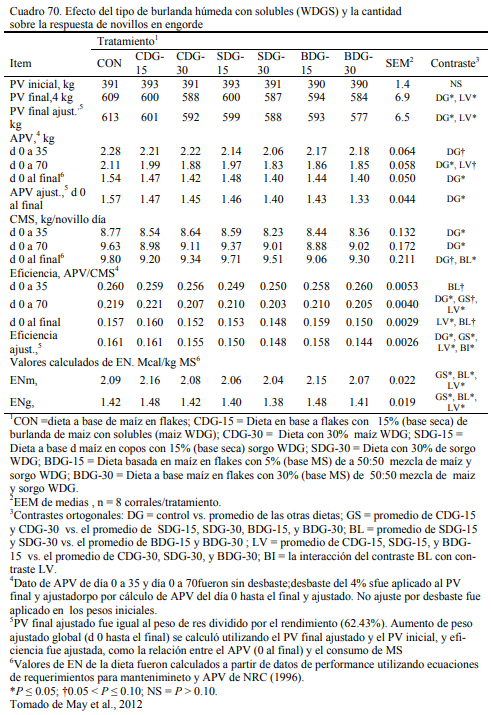
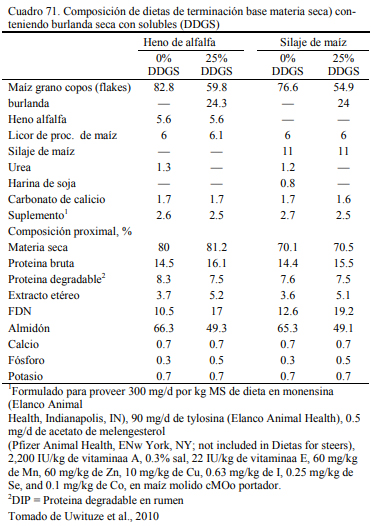
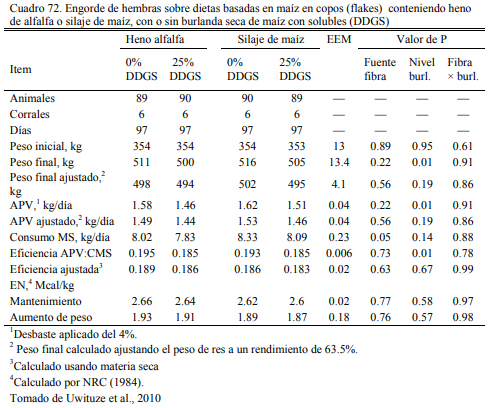
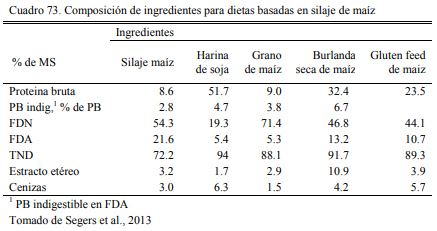
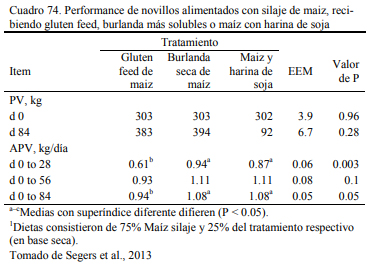
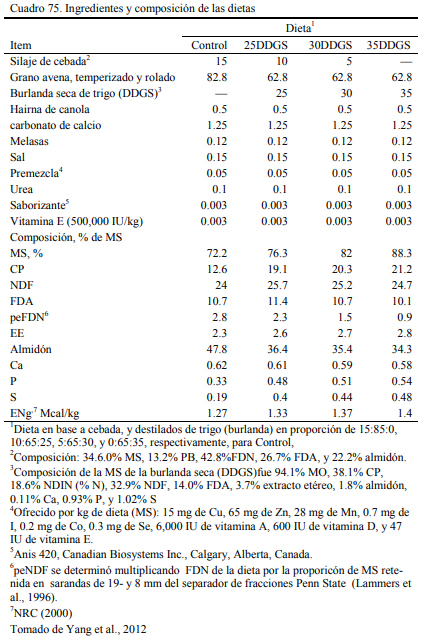
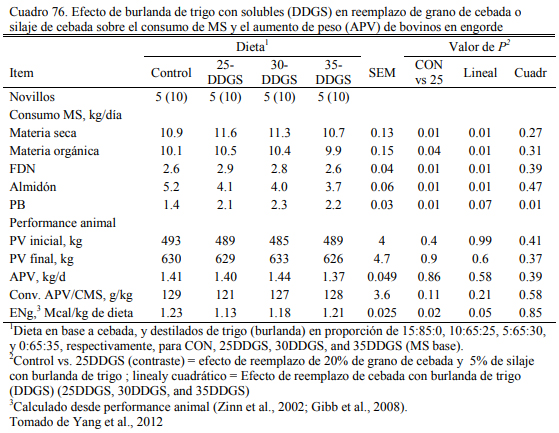
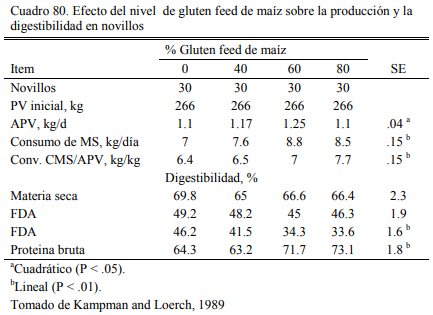
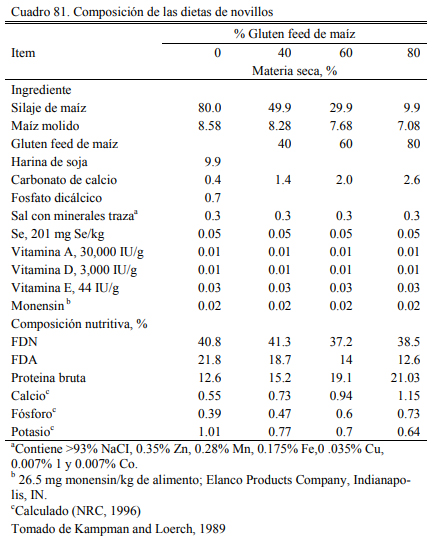
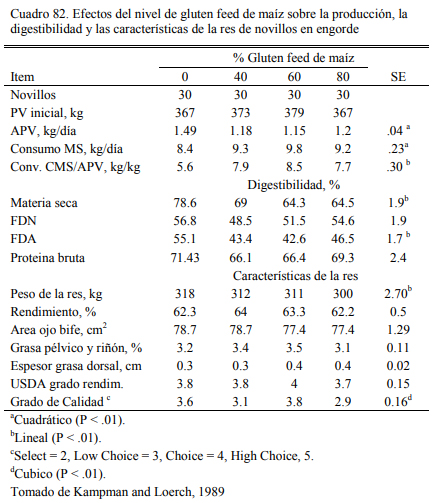
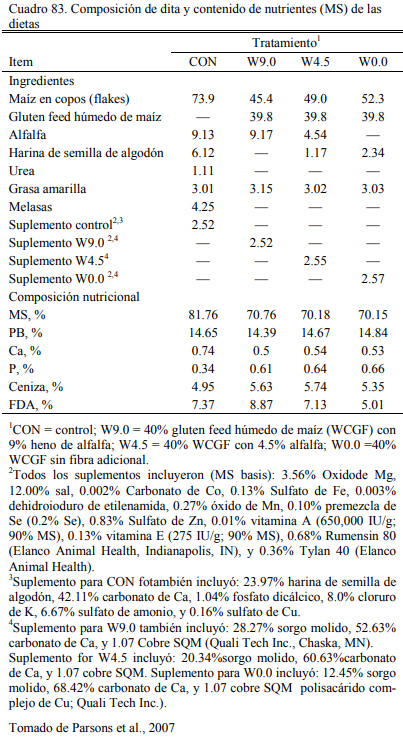
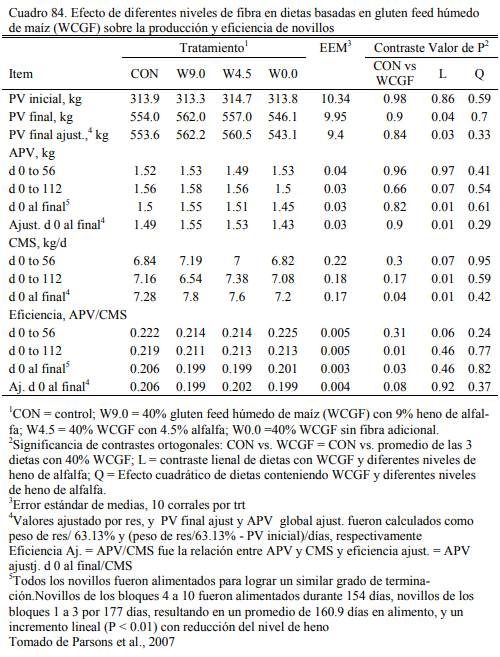
Bibliografía
Ainslie, S. J., D. G. Fox, T. C. Perry, D. J. Ketchen y M. C. Barry. 1993. Predicting amino acid adequacy of diets fed to Holstein steers. J. Anim. Sci.71:1312-1319.
Albro, J. D., D. W. Weber, y T. DelCurto. 1993. Comparison of whole, raw soybeans, extrud- ed soybeans, or soybean meal and barley on digestive characteristics and performance of weaned beef steers consuming mature grass hay. J. Anim. Sci. 71:26–32.
Aldrich, C. G., N. R. Merchen, J. K. Drackley, S. S. Gonzalez, G. C. Fahey, Jr., and L. L. Ber- ger. 1997. The effects of chemical treatment of whole canola seed on lipid and protein digestion by steers. J. Anim. Sci. 75:502–511
Bartle, S.J. y Preston, R.L. 1992. Roughage level and limited maximum intake regimens for feedlot steers. J. Anim. Sci. 70:3293-3303.
Beauchemin, K.A., Yang, W.Z. y Rode, L.M. 2001. Effects of barley grain processing on the site and extent of digestion of feedlot finishing diets. J. Anim. Sci. 7:1925-1936.
Beever, D. E. 1993. Rumen function. In: J. M. Forbes, J. France (Editors). Quantitative Aspets of Ruiminal Digestion and Metabolism. CAB International, Wallingford (UK), pp. 187-215.
Bock, B.J., Brandt, R.T.,Jr., Harmon, D.L., Anderson, S.J., Elliott, J.K., y Avery, T.B. 1991. Mixtures of wheat and high-moisture corn in finishing diets: Feedlot performance and in situ rate of starch digestion in steers. J. Anim. Sci. 69:2703-2710.
Boeto, G.C., O.E. Melo. 1990. Digestión ruminal del almidón en bovinos. I. Efecto del tipo de grano. II. Efecto del tratamiento físico. Informe de beca CONICOR.
Brandt, R. T. y S. J. Anderson. 1990. Supplemental fat source affects feedlot performance and carcass traits of finishing yearling steers and estimated diet net energy value. J. Anim. Sci. 68:2208–2216.
Brandt, R.T., Jr., Anderson, S.J., y Elliott, J.K. 1988. Mixtures of high moisture corn or steam flaked wheat feor finishing cattle. “Cattle Feeders” Day, Rep. Prog. 555, Kans. Agr. Exp. Stn., Garden City, KS. Pg. 32-34.
Brenttheurer, C. 1986. Grain processing effects on starch utilization by ruminants. J. Anim. Sci. 63:1649-1662.
Britton, R.A., y Stock, R.A. 1986. Acidosis, rate of starch digestion and intake. In: Symposium Proceedings: Feed Intake by Beef Cattle. F. N. Owens, Ed. Okla. Agric. Exp. Stn. MP- 121. Pg. 25.
Brown, M.S., Krehbiel, C. R., Galyean, M. L., Remmenga, M. D., Peters, J. P., Hibbard, B., Robinson, J. y Moseley, W. M. 2000a. Evaluation of models of acute and subacute ac- idosis on dry matter intake, ruminal fermentation, blood chemistry, and endocrine pro- files of beef steers. J. Anim. Sci. 78:3155-3168.
Brown, M.S., Krehbiel, C.R., Duff, G.C., Galyean, M.L., Hallford, D.M. y Walker, D.A. 2000b. Effect de degree of corn processing on urinary nitrogen composition, serum metabolite and insulin profiles, and performance by finishing steers. J. Anim. Sci. 78:2464-2474.
Buttrey, E. K., K. H. Jenkins, J. B. Lewis, S. B. Smith, R. K. Miller, T. E. Lawrence, F. T. McCollum III, P. J. Pinedo, N. A. Cole and J. C. MacDonald. 2013. Effects of 35% corn wet distillers grains plus solubles in steam-flaked and dry-rolled corn-based fin- ishing diets on animal performance, carcass characteristics, beef fatty acid composi- tion, and sensory attributes. J. Anim. Sci. 91:1850-1865.
Cardorniga C. y L. D. Satter. 1993. Protein versus energy supplementation of high alfalfa si- lage diets for early lactation cows. J. Dairy Sci. 76:1972-1977.
Castillo, A. R., E. Kebreab, D. E. Beever, J. H. Barbi , J. D. Sutton, H. C. Kirby, and J. France. 2000. J. Anim. Sci.79:240-246.
Castillo, A. R., E. Kebreab, D. E. Beever, J. H. Barbi, J. D. Sutto, H. C. Kirby, and J. France. 2000. The effect of protein supplementation on nitrogen utilization in lactating dairy cows fed grass silage diets. J. Anim. Sci. 79:247-253.
Castro, H., N. Andreo y D. Vottero. 2002. Utilización del silaje de sorgo forrajero azucarado en la terminación a corral de novillos Holando. Rev. Arg. Prod. Anim., 22(1):36.
Chandrashekar, A. y A. W. Kirleis. 1988. Influence of protein on starch gelatinization in sor- ghum. Cereal Chem. 65:457-462.
Cheng, K.J., McAllister, T.A., Popp, J.D., Hristov, A.N., Mir, Z. y Shin, H. T. 1998. A re- view of bloat in feedlot cattle. J. Anim. Sci.78:299-308.
Church, D.C. 1988. The ruminant animal. Digestive physiology and nutrition. Prentice Hall, Englewoods Cliffs, NJ.
Clark, J. H. 1980. Feeding value of high moisture corn for cattle. Pg. 1-11 In: Proc. 41st Min- nesota Nutr. Conf., Minneapolis.
Cooper, R.J., C.T. Milton, T.J. Klopfenstein and D.J. Jordon. 2002a. Effect of corn processing on degradable intake protein requirement of finishing cattle. J. Anim. Sci. 80: 242.
Cooper, R.J., Milton, C.T., Klopfenstein, T.J., Scott,T.L., Wilson, C.B., y Mass, R.A. 2002b. Effect of corn processing on starch digestion and bacterial crude protein flow in finish- ing cattle. J. Anim. Sci. 80:797-804.
Depenbusch, B. E. , E. R. Loe, J. J. Sindt, N. A. Cole, J. J. Higgins and J. S. Drouillard. 2009a. Optimizing use of distillers grains in finishing diets containing steam-flaked corn. J. Anim. Sci. 87:2644-2652.
Depenbusch, B. E., C. M. Coleman, J. J. Higgins and J. S. Drouillard. 2009b. Effects of in- creasing levels of dried corn distillers grains with solubles on growth performance, carcass characteristics, and meat quality of yearling heifers. J. Anim. Sci. 87:2653- 2663
Depetris, G.J., F.J. Santini, Pavan, E., Villarreal, E.L. y. Rearte, D.H. 2003. Efecto del grano de maíz alto en aceite en el sistema de engorde a corral. Rev. Arg. Prod. Anim., 23(1): 57.
Di Marco, O.N. 1993. Crecimiento y respuesta animal. Ed. AAPA.
Duff, G. C., M. L. Galyean, M. E. Branine, D. M. Hallford, M. E. Hubbert, E. Fredrickson y
A. J. Pordomingo. 1990. Effects of continuous versus daily rotational feeding of monensin plus tylosin and lasalocid on serum insulin and growth hormone concentra- tions in beef steers fed a 90% concentrate diet. Proc. Natl. Amer. Soc. Anim. Sci. 41: 55-58
Elizalde, J.C., C.A. Franchone y V.F. Parra. 2003a. Ganancia de peso y eficiencia de conversión en vaquillonas alimentadas a corral con dietas basadas en cebada y afrechillo de trigo. Rev. Arg. Prod. Anim., 23(1): 54.
Elizalde, J.C., C.A. Franchone y V.F. Parra. 2003b. Ganancia de peso y eficiencia de conversión en vacunos alimentados a corral con dietas basadas en granos de maíz entero, cebada entera o aplastada y afrechillo de trigo. Rev. Arg. Prod. Anim. 23(1): 55.
Elizalde, J.C., W. Paul y C. Franchone. 2002. Consumo y ganancia de peso en vacunos alimentados a corral con dietas basadas en grano de maíz entero o molido y con baja inclusión de forraje. Rev. Arg. Prod. Anim., 22(1): 61.
Erickson, G.E., T. J. Klopfenstein, C. T. Milton, D. Brink, M. W. Orth and K. M. Whittet. 2002. Phosphorus requirement of finishing feedlot calves. J. Anim. Sci. 80:1690- 1695.
Erickson, G.E., T.J. Klopfenstein, C. T. Milton, D. Hanson and C. Calkins. 1999. Effect of dietary phosphorus on finishing steer performance, bone status, and carcass maturity. J. Anim. Sci.77:2832-2836.
Eweedah, N., L. Rozsa, J. Gundel, and J. Varhegyi. 1997. Comparison of fullfat soybean, sun- flower seed and protected fat as fat supplements for their effect on the performance of growing-finishing bulls and carcass fatty acid composition. Acta Vet. Hung. 45:151– 163.
Ewing, D.L., Jhonson, D.E. y Rumpler, W.V. 1986. Corn particle passage and size reduction in the rumen of beef steers. J. Anim. Sci.63:1509-1515.
Farran, T. B., G. E. Erickson, T. J. Klopfenstein, C. N. Macken and R. U. Lindquist 2006. Wet corn gluten feed and alfalfa hay levels in dry-rolled corn finishing diets: Effectson fin- ishing performance and feedlot nitrogen mass balance. J. Anim. Sci. 84:1205-1214.
Felice, G., Pordomingo, A. J., Volpi Lagreca, G. y Juan, R. 2013. Raw soybeans in a whole corn diet for feedlot heifers. J. Anim. Sci. (Abstrs.) Indiana T68.
Felton, E.E.D. y M. S. Kerley. 2004. Performance and carcass quality of steers fed whole raw soybeans at increasing inclusion levels. J. Anim. Sci. 82:725–732
Fluharty, F.L., S. C. Loerch, T.B. Turner, S.J. Moeller y G. D. Lowe. 2000. Effects of weaning age and diet on groth and carcass characteristics in steers. J. Anim. Sci. 78:1759-1767.
Fulton, W.R., Klopfenstein, T.J., y Britton, R.A. 1979. Adaptations to high concentrate diet by beef cattle. I Adpatation to corn and wheat diet. J. Anim. Sci.49:775-781.
Galyean, M.L. 1999. Review: Restricted and programmed feeding of beef cattle-definitions, applications and reserach results. Prof. Anim Sci. 15:1-6.
Galyean, M.L., Wagner, D.G. y R. R. Jhonson. 1976. Site y extent of starch digestion in steers fed processed corn rations. J. Anim. Sci. 43:1088-1101.
Ghate, S.R. y Bilansky, W.K. 1981. Preservation of High-Moisture Corn Using Urea. Ameri- can Society of Agricultural Engineers. 24:1047.
Gross, K.L., D.L. Harmon y Avery, T.B. 1988. Net portal nutrient flux in steers fed diets con- taining wheat and sorghum grain alone or in combination. J. Anim. Sci.66:543-551.
Guthrie, M.J., Galyean, M.L., Malcom, K.J., Kloppenburg, J.H. y Wallace, J.D. 1992. Effect of method of corn processing and roughage source on feedlot performance and rumi- nal fermentation in beef steers. Proc. Western Sec., Amer. Soc. Anim. Sci.43:19-22.
Harmon, D.L., Lee, R.W., Milliken, G.A. y Avery, T.B. 1987. Effect of wheat and high- moisture sorghum grain fed singly and in combination on ruminal fermentation, solid and liquid flow, site and extent of digestion and feeding performance of cattle.J. Anim. Sci. 64: 897-906.
Hejazi, S., Fluharty, F.L., Perley, J.E., Loerch, S.C. y Lowe, G.D. 1999. Effect of corn pro- cessing and dietary fiber source on feedlot performance, visceral organ weight, diet digestibility and nitrogen metabolism in lambs. J. Anim. Sci. 77:507-515.
Herrera-Saldana, R. E., Huber, J. T. y Poore, M. H. 1990. Dry matter, crude protein and starch degradability of five cereal grains. J. Dairy Sci. 73:2386-2393.
Herrera-Saldana, R., R. Gomez Alarcón, M. Torabi y J. T. Huber. 1990. Influence on synchro- nizing protein and starch degradation in the rumen of nutrient utilization and microbial protein synthesis. J. Dairy Sci. 73:142-148.
Hibberd, C.A., Wagner, D.G., Schemm, R.L., Mitchell, Jr., E..D., Weibel, D.E. y Hintz, R.L. 1982. Digestibility characteristics of isolated starch from sorghum and corn grain. J. Anim. Sci. 55:1490-1497.
Hicks, R.B., Owens, F.N., Gill, D.R., Martin, J.J. y Strasia, C.A. 1996. Effects of controlled fed intake on performance and carcass characteristics of feedlot steers and heifers. J. Anim. Sci.68:233-244.
Hill, W.J., Secrist, D.S., Owens, F. N., Strasia, C.A, Gill, D.R., Basalan, M. y Johnson, A.B. 1996. Effects of trace mineral supplements on performance of feedlot steers. Okla. Agric. Exp. Sta.Misc. Pub. P-951, 153-163.
Hoover, W. H. y S.R. Stokes. 1993. Balancing carbohydrates and proteins for optimun micro- bial yield. J. Dairy Sci. 74:3630-3644.
Huck, G.L., Kreikemeier, K.K., Kuhl, G.L., Eck T.P. y Bolsen, K.K. 1998. Effects of feeding combinations of steam-flaked grain sorghum and steam-flaked, high-moisture, or dry rolled corn on growth performance and carcass characteristics in feedlot cattle. J. Anim. Sci. 76:2984-2990.
Huerta-Leidenz, N. O., H. R. Cross, D. K. Lunt, L. S. Pelton, J. W. Savell, y S. B. Smith. 1991. Growth, carcass traits, and fatty acid profiles of adipose tissues from steers fed whole cottonseed. J. Anim. Sci.69:3665–3672.
Huntington, G.B. 1997. Starch utilization for ruminants: From basics to bunk. J. Anim. Sci.75:852-867.
Juan, N., A.J. Pordomingo y R. Jouli. 1997. Engorde de vaquillonas a corral con silaje de maíz y heno de alfalfa. Rev. Arg. Prod. Anim.17(1):280.
Juan, N.A. Pordomingo, A.J. y Jouli, R.R. 1998b. Reemplazo de silaje de grano húmedo de maíz por grano húmedo de sorgo en dietas para engorde. Rev. Arg. Prod. Anim. 18(1):50.
Juan, N.A., Pordomingo, A.J., Velilla, S.M. y Jouli R.R. 1998a. Utilización de grano húmedo de sorgo conservado con urea para engorde de vaquillonas. Rev. Arg. Prod. Anim. 18(1):48.
Kamoman. K. A. y Loerch, S. C. 1989. Effects of dry corn gluten feed on feedlot cattle per- frmance and fiber digestibility. J. Anim. Sci. 67:501-512.
Kauffman, W., Hageneister, H., Dirksen, G. 1980. Adaptation changes in dietary composition, level and frequency of feeding. In: Digestive Physiology and Metabolism in Rumi- nants Y. Ruckebusch y Thivend, P., Eds. AVI Publishing, Wesport, CT, USA. p. 587- 595.
Kaufmann, W., 1976. Influence of the composition of the ration and the feeding frequency on pH regulation in the rumen and on feed intake in ruminant livestock. Prot. Sci. 3:103- 108.
Kennelly, J. J., Dalton, D. L. y HA, J. K. 1988. Digestion y utilization of high moisture barley by lactating dairy cows. J. Dairy Sci. 71:1259-1266.
Kent, F., Pordomingo, A.J. y Jouli, R. 2011. Efecto del tipo de sorgo sobre la performance animals en dietas basadas en silaje. Rev. Arg. Prod. Anim. 31(1):313.
Kincaid, R. 1988. Macro-elements for ruminants. In: The Ruminant Animal. Digestive physi- ology and nutrition. D. C. Church (Ed.). Prentice Hall. Englewood Cliffs, NY,
Knaus, W. F., D. H. Beermann, T. F. Robinson, D. G. Fox y K. D. Finnerty. 1998. Effects of dietary mixture of meat and bone meal, feather meal, blood meal, and fish meal on ni- trogen utilization in finishing Holstein steers. J. Anim. Sci. 76:1481-1487.
Koenig, K. M. and K. A. Beauchemin. 2013a. Nitrogen metabolism and route of excretion in beef feedlot cattle fed barley-based finishing diets varying in protein concentration and rumen degradability. J. Anim. Sci. 91:2310-2320.
Koenig, K. M. and K. A. Beauchemin. 2013b. Nitrogen metabolism and route of excretion in beef feedlot cattle fed barley-based backgrounding diets varying in protein concentra- tion and rumen degradability. J. Anim. Sci. 91:2295-2309.
Kreikemeier, K.K., Harmon, D.L., Brandt, R.T., Jr., Nagaraja, T.G. y Cochran, R.C. 1990. Steam-rolled wheat diets for finishing cattle: Effects of dietary roughage and feed in- take on finishing steer performance and ruminal metabolism. J. Anim. Sci. 68:2130- 2141.
Kreikemeier, K.K., Stock, R.A., Brink, D.R. y Britton, R.A. 1987. Feeding combinations of dry corn and wheat to finishing lambs and cattle. J. Anim. Sci.65:1647-1651.
Lee, J., P. M. Harris, B. R. Sinclair y B. P Treloar. 1992. The effect of condensed tannin con- taining diets on whole body amino acid utilization in Rommey sheep: Consequences for wool growth. Proc. N. Z. Soc. Anim. Prod. 52:243-245.
Lee, R. W., Galyean, M. L. y Lofgreen, G. P. 1982. Effects of mixing whole, shelled and steam-flaked corn in finishing diets on feedlot performance and site and extent of di- gestion in beef steers. J. Anim. Sci. 55:475-485.
Loerch, S.C. y Berger, L.L. 1980. Protein solubility and in-situ protein disappearance as af- fected by pH and ration energy level. J. Anim. Sci. 50(1):377-382.
Loerch, S.C. y Fluharty, F.L. 1998a. Effects of corn processing, dietary roughage level, and timing of roughage inclusion on performance of feedlot steers. J. Anim. Sci. 76:681- 685.
Loerch, S.C., y Fluharty F.L. 1998b. Effects of programming intake on performance and car- cass characteristics of feedlot cattle. J. Anim. Sci. 76:371-377.
Lyle, R.R., Johnson, R.R., Wilhite, J.V. y Backus, W.R. 1981. Ruminal characteristics in steers as affected by adaptation from forage to all concentrate diets. J. Anim. Sci. 53:1383-.
Mabuku, O., Hatendi, P.R. y Medlinah, M. 1996. Effect of feeding different mixtures of whole to milled maize grain in a complete diet on feedlot performance of steers. Zimbabwe J. Agric. Res. 34:87-95.
Macken, C. N., G. E. Erickson, T. J. Klopfenstein and R. A. Stock Effects of concentration and composition of wet corn gluten feed in steam-flaked corn-based finishing diets. 2004. J. Anim. Sci.82:2718-2723.
MacRae, J.E. y Amstrong, D.G. 1968. Enzime method for determination of alpha-linked glu- cose polimers in biological materials. J. Sci. Food Agric. 19:578.
Mader, T.L., Poppert, G.L. y Stock, R.A. 1993. Evaluation of alfalfa type as a roughage source in feedlot adaptation and finishing diets containing different corn types. Animal Feed Sci. and Tech. 42:109-119.
Maresca, S., F.J. Santini, E. Pavan, Elizalde J.C. y G. Eyherabide. 2003. Efecto del nivel de forraje en la dieta sobre la utilización del grano de maíz entero en bovinos de diferentes edades. Rev. Arg. Prod. Anim., 23(1):38.
Maresca, S., Santini, F.J. y Paván, E. 2002. Comportamiento productivo de terneras alimenta- das a corral con grano de maíz entero y partido. Rev. Arg. Prod. Anim. 22:163-168.
May, M. L., J. C. DeClerck, M. J. Quinn, N. DiLorenzo, J. Leibovich, D. R. Smith, K. E. Hales and M. L. Galyean. 2010. Corn or sorghum wet distillers grains with solubles in combination with steam-flaked corn: Feedlot cattle performance, carcass characteris- tics, and apparent total tract digestibility. J. Anim. Sci.88:2433-2443.
McCollough, R. L. y Brent, B. E. 1972. Digestibility of eight hybrid sorghum grains and three hybrid corns. Kansas Agric. Exp. Sta. Bull. 557:27.
McNeill, J.W., Potter, G.D. y Riggs, J. K. 1976. Ruminal and pos-ruminal carbohydrate utili- zation in steers fed processed sorghum grain. J. Anim. Sci. 33:1371-1388.
Meissner, et al. 1992. Effect of level and degradation of dietary protein on performance of feedlot steers. J. Anim. Sci. 22:4.
Meyer, N. F, G. E. Erickson, T. J. Klopfenstein, J. R. Benton, M. K. Luebbe and S. B. Laudert. 2013. Effects of monensin and tylosin in finishing diets containing corn wet distillers grains with solubles with differing corn processing methods. J Anim. Sci 91:2219- 2228.
Morgan, C.A. y Campling, R.C. 1978. Digestibility of whole barley and oat grains by cattle of different ages. Anim. Prod. 27:323-329.
Mueller, J. C., Blalock, H. M. y Pritchard, R. H. 2011. Use of soybean hulls as a replacement for dry rolled corn in beef cattle feedlot receiving diets. J. Anim. Sci. 89:4142-4150.
Murphy, T.A. y Loerch, S.C. 1994. Effects of restricted feeding of growing steers on perfor- mance, carcass, characteristics, and composition. J. Anim. Sci. 72:2497-2507.
Murphy, T.A., Fluharty, F.L. y Loerch, S.C. 1994a. The influence of intake level and corn processing on digestibility and ruminal metabolism in steers fes all-concentrate diets. J. Anim. Sci. 72:1608-1615.
Murphy, T.A., Loerch, S.C. y Dehority, B.A. 1994b. The influence of restricted feeding on site and extent of digestion and flow of nitrogenous compounds to the duodenum in steers. J. Anim. Sci. 72:2487-2496.
Nocek, J. E. y Tamminga, S. 1991. Site of digestion of starch in the gastrointestinal tract of dairy cows and its effects on milk yield and composition. J. Dairy Sci. 74:3598-3629.
NRC, 2000. Nutrient Requirement for Beef Cattle (8th) Ed. National Academy Press. Wash- ington, DC.
NRC. 1996. Nutrient requirements of beef cattle (7th Ed.). National Academy Press, Washing- ton, D.C.
Orskov, E. R., Mayes R. W. y Penn, A. 1971. The capacity for thew removal of glucose from the small intestine by mature sheep. Proc. Nutr. Soc. 30:43A.
Owens, F. N., Secrist, D., Hill, J., Gill, D. 1996. A new look at acidosis. In: Proc. Southwest Nutrition and Management Conf. University of Arizona, Tucson. Pp 1-16.
Owens, F.N., Zin, R.A. y Kim, Y.K. 1986. Limits to starch digestion in the ruminant small intestine. J. Anim. Sci. 63:1634-1652.
Owens, F.N., Secrist, D.S., Hill, W.J. y Gill, D.R. 1997. The effect of grain source and grain processing on performance of feedlot cattle: A review. J. Anim. Sci. 75:868-879.
Parra, V.F., J.C. Elizalde y G.A. Duarte. 2002. Resultados de engordes a corral de vacunos en diferentes sistemas de producción de carne. Rev. Arg. Prod. Anim., 22(1): 60.
Perry, T.W. 1976. The feeding value of high-moisture grains for beef cattle. High Moisture Grains Symp. Oklahoma State Univ., Stillwater.
Petit, H. L. y G. F. Tremblay, 1995. Ruminal fermentation and digestion in lactating cows fed grass silage with protein and energy supplements. J. Dairy Sci. 78:342-352.
Philippeau, C., C. Martin y B. Michalet-Doreau. 1999a. Influence of grain source on ruminal characteristics and rate, site, and extent of digestion in beef steers. J. Anim. Sci. 77:1587-1596.
Philippeau, C., F. Le Deschault de Monredon, y B. Michalet-Doreau. 1999b. Relationships between ruminal starch degradation and the physical characteristics of corn grain. J. Anim. Sci. 77:238-243.
Pordomingo, A. J., Pordomingo, A. B. y Volpi Lagreca, G .2007a. Efecto del uso de cebada en dietas de engorde en confinamiento de terneras para carne. Rev. Arg. Prod. Anim. 27(1):80-81.
Pordomingo, A. J., Pordomingo, A. B., Miranda, A. y Juan, N. 2007b. Efecto del uso de afre- chillo de trigo en dieta basada en grano entero en engorde de terneros. Rev. Arg. Prod. Anim. 27(1):85-86.
Pordomingo, A. J., Kent, F., Jouli, R. y Felice, G. 2012. Sustitución de silaje de maiz por grano de maiz en dietas de feedlot. Rev. Arg. Prod. Anim. 32(1)344.
Pordomingo, A. J., Volpi Lagreca, G., Stefanazzi, I.N. y Pordomingo, A.B. 2007c. Inclusión de taninos, monensina y soja cruda en dietas de grano entero en engorde de vaquillo- nas a corral. Rev. Arg. Prod. Anim. 27(1):81-83.
Pordomingo, A.J. 2002. La edad al destete, la fuente y el nivel de fibra en la dieta del ternero de destete precoz. Rev. Arg. Prod. Anim. 22:1-13
Pordomingo, A.J. 1990. Effects of daily and weekly rotations of lasalocid and monensin plus tylosin compared with continuous feeding of ionophores on ruminal fermentation in beef steers fed a 90% concentrate diet. PhD Dissertation, New Mexico State Univ. USA pp. 1-12.
Pordomingo, A.J. y G. Volpi Lagreca. 2004. Efectos de la concentración vitamínica y mineral de la dieta, el enriquecimiento con metionina y la presentación del grano de maíz so- bre el crecimiento de terneros destetados precozmente y alimentados a corral con die- tas de alta energía. Rev. Arg. Prod. Anim. 24(1):65.
Pordomingo, A.J. y Juan, N. A. 2000. Grano húmedo de sorgo conservado con urea: Efecto del partido sobre su potencial de engorde. Rev. Arg. Prod. Anim. 20(1):91.
Pordomingo, A.J., Jonas, O., Adra, M., Santucho, G., Juan, N.A. y Azcárate, M.P. 2002a. Evaluación de dietas basadas en grano entero, sin fibra larga, en engorde a corral. RIA 31:1-22.
Pordomingo, A.J., Galyean, M. E., Branine, M. E. and Freeman, A.S. 1999a. Effects of daily and weekly rotations of lasalocid and monensin plus tylosin compared with continuous feeding of each ionophore on daily dry matter intake and digesta kinetics. Rev. Arg. Prod. Anim. 19:383 – 390.
Pordomingo, A.J., Galyean, M. E., Branine, M. E. y Freeman, A.S. 1999b. Effects of daily and weekly rotations of lasalocid and monensin plus tylosin compared with continuous feeding of ionophores on ruminal fermentation. Rev. Arg. Prod. Anim.19:362-382
Pordomingo, A.J., Jonas, O., Orroño, R., Juan, N.A. y Azcárate, M.P. 2002c. Efecto de la hu- medad del grano y el agregado de urea sobre la conservación alcalina del maiz. Rev. Arg. Prod. Anim. 22(1):84
Pordomingo, A.J., Juan, N.A. y Azcarate, P. 2002b. Grano de maíz húmedo conservado con urea en dietas de engorde a corral. Rev. Arg. Prod. Anim. 22:101-113
Pordomingo, A.J., N.A. Juan y M.P. Azcarate. 2003. .Effect of condensed-tannins addition to a corn-sunflower meal based feedlot diet. J. Anim. Sci.81(1):215.
Pordomingo, A.J., Pordomingo, A.B. y Lernoud, P. 2011. Sustitución de maíz aplastado por cebada aplastada en dietas para engorde de novillos a corral. Rev. Arg. Prod. Anim. 31(1):314.
Pordomingo, A.J., Volpi Lagreca, G., Orienti, W. y Welsh, R. 2004. Evaluación del agregado de taninos en dietas de distinto nivel energético en vaquillonas para carne. Rev. Arg. Prod. Anim. 24(1):67.
Raun, A.P., Cooley, C.O., Potter, E.L., Rathemacher, R.P. y Richardson, L.F. 1976. Effect of monensin on feed efficiency of feedlot cattle. J. Anim. Sci. 43-670-
Reinhardt, C.D., Brandt, R.T. Jr., Eck, T.P. y Titgemeyer, E.C. 1998. Performance, digestion and mastication efficiency of Holstein steers fed whole or processed corn in limit- or full-fed growing-finishing systems. J. Anim. Sci. 76:1778-1788.
Rooney, L. W. y Pflugfelder, R. L. 1986. Factors affecting starch digestibility with special emphasis on sorghum and corn, J. Anim. Sci. 63:1607-1609.
Rossi, J.E., Loerch, S.C. y Fluharty, F.L. 2000. Effects of crude protein concentration in diets of feedlot steers fed to achieve stepwise increases in rate of gain. J. Anim. Sci. 78:3036-3044.
Russell, J R., Young A.W. y Jorgensen, N. A. 1981. Effect of dietary cornstarch intake on pancreatic amylase, intestinal maltase and pH in cattle. J. Anim. Sci. 52:1177-1196.
Russell, J. B y Rychlik. 2001. Factors that alter rumen microbiology. Science. 292:2679- 2692.
Russell, W. y Schmidt, S.P. 1993. Preservation of High-Moisture Sorghum Grain with Urea and Evaluation in Cattle Feeding. West Virginia University & Auburn University. 18- th Biennial Grain Sorghum Research and Utilization Conference, 9 pp.
Russell, W., Lin, J.C.M., Thomas, E.E. y Mora, E. C. 1988. Preservation of high-moisture milo with urea: grain properties and animal acceptability. J. Anim. Sci.66:2131.
Sainz, M.L., De la Torre, F., y Oltjen, J. W. 1995. Compensatory growth and carcass quality in growth-restricted and reved beef steers. J. Anim. Sci. 73:2971-2979.
Santini, F. J. y E. Pavan. 2002. Alimentación a corral. Del silaje al grano de maíz. IDIA XXI, Nº2, 30-34.
Santini, F.J. y Di Marco, O.N. 1983. Monensina. Modo de acción y su efecto sobre el compor- tamiento productivo del animal. Rev. Arg. Prod. Anim. 3:345-
Santini, F.J., Villarreal, E.L. Pavan, E. y Grigera, J.M. 2003. Características productivas de novillos de diferente tamaño estructural engordados a corral con dietas de diferente concentración energética. Rev. Arg. Prod. Anim., 23(1): 48.
Schoonmaker, J. P., A. H. Trenkle and D. C. Beitz. 2010. Effect of feeding wet distillers grains on performance, marbling deposition, and fatty acid content of beef from steers fed low- or high-forage diets. J. Anim. Sci. 88:3657-3665.
Secrist, D.S., Hill, W.J., Owens, F.N., Gill, D.R. y Welty, S.D. 1996a. Rolled or whole corn for feedlot steers being limit-or ad libitum-fed. Okla. Agric. Exp. Sta.Misc. Pub.. P- 951,173-180.
Secrist, D.S., Owens, F.N., Hill, W.J. y Welty, S.D. 1996b. Rolled versus whole corn: Effects on ruminal fermentation of feedlot steers. Okla. Agric. Exp. Sta.Misc. Pub. P-951, 181-188.
Segers, J. R., A. M. Stelzleni, T. D. Pringle, M. A. Froetschel, C. L. Ross and R. L. Stewart, Jr. 2013. Use of corn gluten feed and dried distillers grains plus solubles as a replacement for soybean meal and corn for supplementation in a corn silage-based stocker system. J. Anim. Sci. 91:950-956.
Sinclair, L. A., P. C. Garnsworthy, J. R. Newbold y P. J. Buttery. 1995. Effects of synchroniz- ing the rate of dietary energy and nitrogen release in diets with a similar carbohydrate composition on the rumen fermentation and microbial protein synthesis in sheep. J. Agric. Sci. 124:463-472.
Sip, M.L. y Pritchard, R. H. 1991. Nitrogen utilization by ruminants during restricted intake of high-concentrate diets. J. Anim. Sci. 69:2655-2662.
Smith, S. B., y J. D. Crouse. 1984. Relative contributions of acetate, lactate and glucose to lipogenisis in bovine intramuscular and subcutaneous adipose tissue. J. Nutr. 114:792– 800.
Spears, J.W., Hatfield, E.E. y Clark, J.H. 1980. Influence of formaldehyde treatment of soy- bean meal on performance of growing steers and protein availability in the chick. J. Anim. Sci. 62:521-526.
Steen, R.W.J. y D.J. Kilpatrick. 2000. The effects of the ratio of grass silage to concentrates in the diet and restricted dry matter intake on the performance and carcass composition of beef cattle Livestock Prod. Sci.62:181-192.
Stern, M.D., G. A. Varga, J. H. Clark, J.L. Firkins, J.T. Huber y D. L. Palmquist. 1994. Evalu- ation of chemical and physical properties of feeds that affect protein metabolism in the rumen. J. Dairy Sci. 69:2734-2739.
Stock R. A., Brink, D. R., Britton R. A., Goedeken, F. K., Sindt, M. H. Kkreikemeier, K. K., Bauer, M. L. y Smith, K. K. 1987a. Feeding combinations of high moiture corn and dry-rolled grain sorghum to finishing steers. J. Anim. Sci. 65:290-302.
Stock, R., N. Merchen, T. Klopfenstein y M. Poos. 1989. Feeding value of slowly degraded proteins. J. Anim. Sci. 53:1109-1119.
Stock, R.A., Brink, D. R., Brandt, R.T., Merrill, J.K. y Smith, K.K. 1987b. Feeding combina- tions of high moisture corn and dry corn to finishing cattle. J. Anim. Sci. 65:282-289.
Stock, R.A., Klopfenstein, T. y Shain, D. 1995. Feed intake variation. Okla. Agric. Exp. Sta.Misc. Publ. P-942:56-59.
Storry, J. E. and J. D. Sutton. 1969. The effect of change from low roughage to high roughage diets on rumen fermentation, blood composition and milk fat secretion in the cow. British J. Nutr. 23:511-518.
Streeter, M.N., Wagner, D.G., Owens, F.N. y Hibberd, C.A. 1989. Combinations of high- moisture harvested sorghum grain and dry-rolled corn: Effects on site and extent of digestion in beef heifers. J. Anim. Sci. 67:1623-1633.
Surber, L.M.N. y Bowman, J.G.P. 1998. Monensin effects on digestion of corn or barley high- concentrate diets. J. Anim. Sci. 76:1945-1954.
Swanson, K.C., Richards, C.J. y Harmon, D.L. 2002. Influence of abomasal infusion of glu- cose or partially hydrolyzed starch on pancreatic exocrine secretion in beef steers. J. Anim. Sci.8:1112-1116.
Swingle, R.S., Eck, T.P., Theurer, C.B. De la Plata, M., Poore, M. H. y Moore, J. A. 1999. Flake density of steam-processed sorghum grain alters peformance and sites of digest- ibility by growing-finishing steers. J. Anim. Sci. 77:1055-1065.
Swingle, S. 1995. Effect of roughage level and type on intake and performance of feedlot cat- tle. Okla. Agric. Exp. Sta. Misc. Pub. P-942:257.
Theurer, C.B. 1986. Grain processing effects on starch utilization by ruminants. J. Anim. Sci.63:1624-1643.
Theurer, C.B., Swingle, R.S., Wanderley, R.C., Kattnig, R.M., Urias, A. and Ghenniwa, G. 1999. Sorghum grain flake and source of roughage in feedlot cattle diets. J. Anim. Sci.77:1066-1073.
Titgemeyer, E. C., N. R. Merchen, y L. L. Berger. 1989. Evaluation of soybean meal, corn gluten meal, blood meal and fish meal as sources of nitrogen and amino acids disap- pearing from the small intestine of steers. J. Anim. Sci. 67:262-275.
Turgeon, O.A., Brink, D.R. y Briton, R.A. 1988. Corn particle size mixture roughage level and starch utilization in finishing steer diets. J. Anim. Sci. 57:739.
Utley, P. R., R. E. Hellwig, J. L. Butler and W. C. McCormick 1973. Comparison of Un- ground, Ground and Pelleted Peanut Hulls as Roughage Sources in Steer Finishing Di- ets. J. Anim. Sci. 37:608-611.
Uwituze, S., G. L. Parsons, M. K. Shelor, B. E. Depenbusch, K. K. Karges, M. L. Gibson, C. D. Reinhardt, J. J. Higgins and J. S. Drouillard. 2010. Evaluation of dried distillers grains and roughage source in steam-flaked corn finishing diets. J. Anim. Sci. 88:258- 274.
Vander Pol, K. J., M. K. Luebbe, G. I. Crawford, G. E. Erickson,3 and T. J. Klopfenstein. 2009. Performance and digestibility characteristics of finishing diets containing distill- ers grains, composites of corn processing coproducts, or supplemental corn oil. J. Anim. Sci. 87:639–652
Volpi Lagreca, G., Alende , M., Pordomingo, A. J. 2010. Efecto de la sustitución de harina de girasol por urea en dietas basadas en silaje de planta entera de sorgo en engorde de terminación de novillos. Rev. Arg. Prod. Anim. 30(1): 491-492.
Waldo, D. R. 1973. Extent and partition of cereal grains starch digestion in ruminants. J. Anim. Sci.37:1062-1083.
Waldo, D.R. 1976. Effects of buffers on ruminal acids. In: Buffers in ruminants. J. Anim. Sci.37:1062-1068.
Wang, Y., Creer, D. y McAllister, T.A. 2003. Effects of moisture, roller setting, and saponin- based surfactant on barley processing, ruminal degradation of barley, and growth per- formance by feedlot steers. J. Anim. Sci. 81:2145-2154.
Wessels, R. H., y Titgemeyer, E.C. 1997. Protein requirements of growing steers limit-fed corn-based diets. J. Anim. Sci. 75:3278-3286.
Wierenga, K. T., T. A. McAllister, D. J. Gibb, A. V. Chaves, E. K. Okine, K. A. Beauchemin and M. Oba 2010. Evaluation of triticale dried distillers grains with solubles as a sub- stitute for barley grain and barley silage in feedlot finishing diets, J. Anim. Sci. 88:3018-3029.
Yang, W. Z., Y. L. Li, T. A. McAllister,* J. J. McKinnon and K. A. Beauchemin. 2012. Wheat distillers grains in feedlot cattle diets: Feeding behavior, 1,2 growth perfor- mance, carcass characteristics, and blood metabolites. J. Anim. Sci.90:1301–1310.
Zin, R.A. y Owens, F.N. 1983. Influence of feed intake level on site of digestion in steers fen a high concentrate diet. J. Anim. Sci. 56:471.
Autor/es

Ingeniero Agrónomo en la Facultad de Agronomía de la Universidad Nacional de La Pampa. Realizó sus estudios de posgrado en New Mexico State University, Las Cruces, New Mexico, USA obteniendo los grados de Master in Animal Science (Animal Nutrition) (1987-1989) y Doctor of Philosophy (PhD) en Ciencia Animal (1988-1991)